

Glutaredoxin GRXS13 plays a key role in protection against photooxidative stress in Arabidopsis
- PMID: 21963612
- PMCID: PMC3245481
- DOI: 10.1093/jxb/err301
Glutaredoxin GRXS13 plays a key role in protection against photooxidative stress in Arabidopsis
Abstract
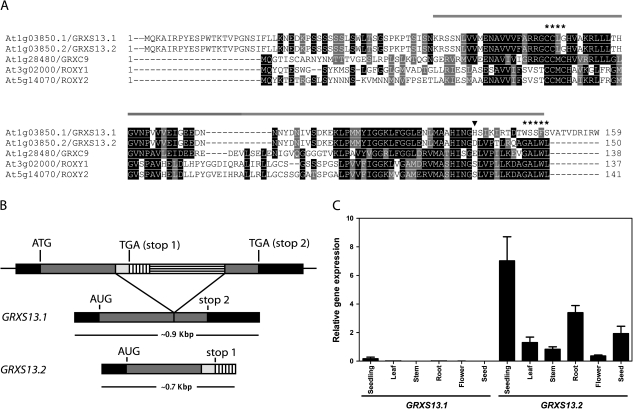
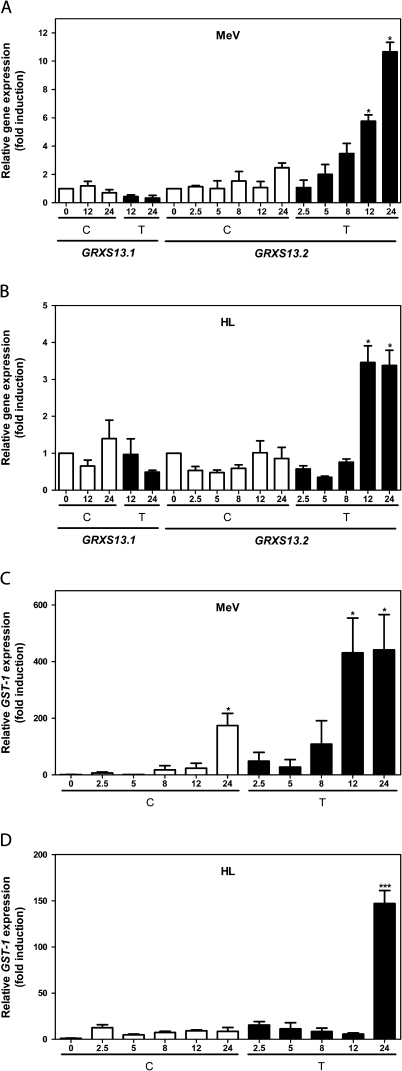
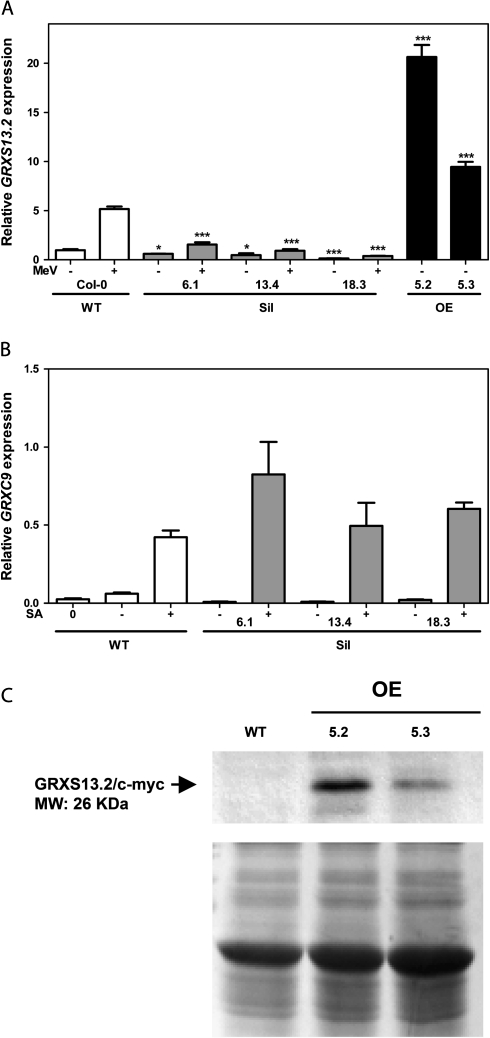
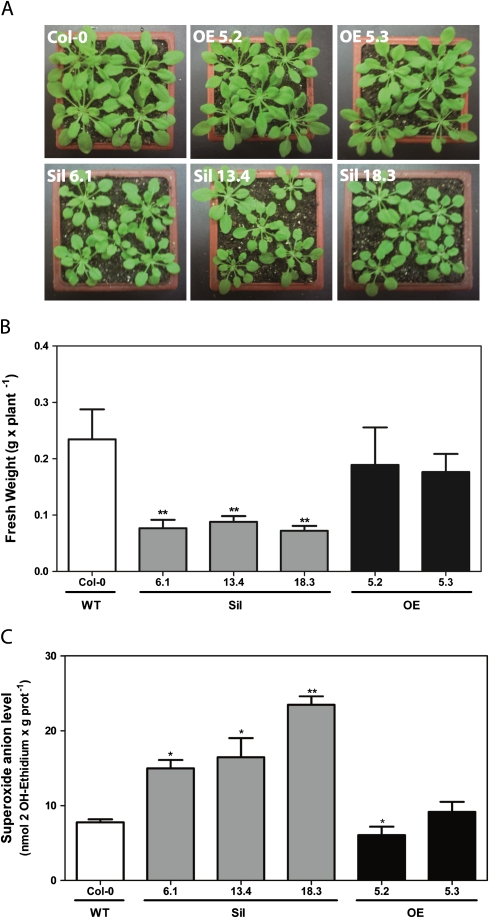
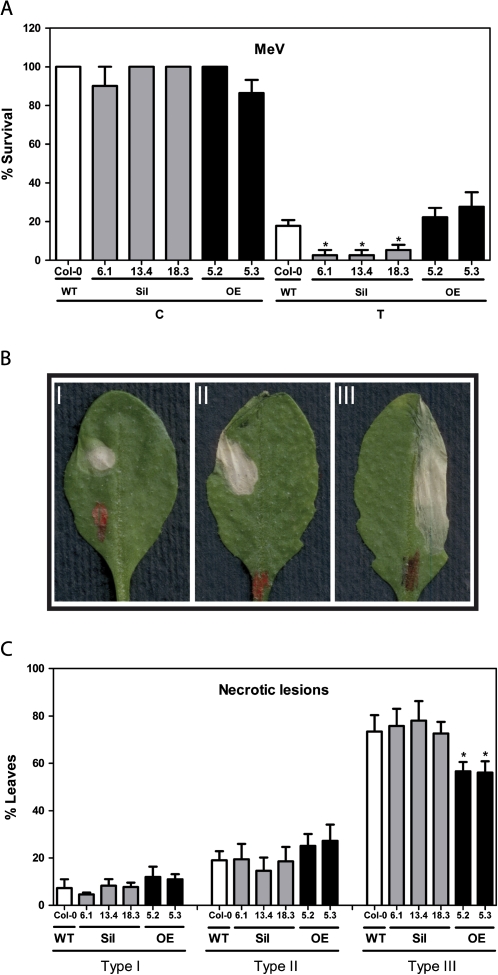
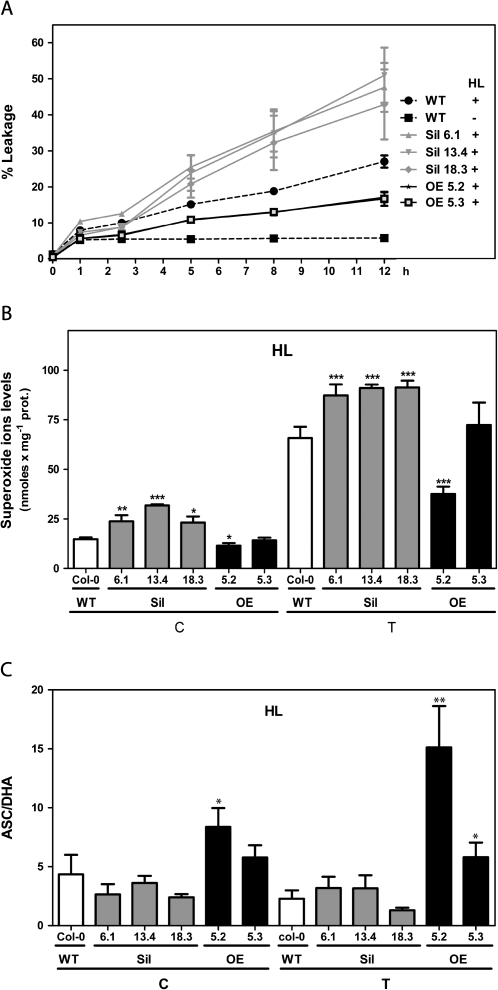
Glutaredoxins (GRXs) belong to the antioxidant and signalling network involved in the cellular response to oxidative stress in bacterial and eukaryotic cells. In spite of the high number of GRX genes in plant genomes, the biological functions and physiological roles of most of them remain unknown. Here the functional characterization of the Arabidopsis GRXS13 gene (At1g03850), that codes for two CC-type GRX isoforms, is reported. The transcript variant coding for the GRXS13.2 isoform is predominantly expressed under basal conditions and is the isoform that is induced by photooxidative stress. Transgenic lines where the GRXS13 gene has been knocked down show increased basal levels of superoxide radicals and reduced plant growth. These lines also display reduced tolerance to methyl viologen (MeV) and high light (HL) treatments, both conditions of photooxidative stress characterized by increased production of superoxide ions. Consistently, lines overexpressing the GRXS13.2 variant show reduced MeV- and HL-induced damage. Alterations in GRXS13 expression also affect superoxide levels and the ascorbate/dehydroascorbate ratio after HL-induced stress. These results indicate that GRXS13 gene expression is critical for limiting basal and photooxidative stress-induced reactive oxygen species (ROS) production. Together, these results place GRXS13.2 as a member of the ROS-scavenging/antioxidant network that shows a particularly low functional redundancy in the Arabidopsis GRX family.
Figures
Similar articles
-
A Rice CPYC-Type Glutaredoxin OsGRX20 in Protection against Bacterial Blight, Methyl Viologen and Salt Stresses.Front Plant Sci. 2018 Feb 9;9:111. doi: 10.3389/fpls.2018.00111. eCollection 2018. Front Plant Sci. 2018. PMID: 29479359 Free PMC article.
-
Over-expression of chickpea glutaredoxin (CaGrx) provides tolerance to heavy metals by reducing metal accumulation and improved physiological and antioxidant defence system.Ecotoxicol Environ Saf. 2020 Apr 1;192:110252. doi: 10.1016/j.ecoenv.2020.110252. Epub 2020 Jan 31. Ecotoxicol Environ Saf. 2020. PMID: 32014725
-
AtGRXcp, an Arabidopsis chloroplastic glutaredoxin, is critical for protection against protein oxidative damage.J Biol Chem. 2006 Sep 8;281(36):26280-8. doi: 10.1074/jbc.M601354200. Epub 2006 Jul 7. J Biol Chem. 2006. PMID: 16829529
-
Plant-specific CC-type glutaredoxins: functions in developmental processes and stress responses.Biol Chem. 2015 May;396(5):495-509. doi: 10.1515/hsz-2014-0300. Biol Chem. 2015. PMID: 25781542 Review.
-
Functions of heat shock transcription factors involved in response to photooxidative stresses in Arabidopsis.Biosci Biotechnol Biochem. 2016 Jul;80(7):1254-63. doi: 10.1080/09168451.2016.1176515. Epub 2016 Apr 20. Biosci Biotechnol Biochem. 2016. PMID: 27095030 Review.
Cited by
-
Chickpea glutaredoxin (CaGrx) gene mitigates drought and salinity stress by modulating the physiological performance and antioxidant defense mechanisms.Physiol Mol Biol Plants. 2021 May;27(5):923-944. doi: 10.1007/s12298-021-00999-z. Epub 2021 May 6. Physiol Mol Biol Plants. 2021. PMID: 34092945 Free PMC article.
-
Redox Modulation Matters: Emerging Functions for Glutaredoxins in Plant Development and Stress Responses.Plants (Basel). 2014 Nov 25;3(4):559-82. doi: 10.3390/plants3040559. Plants (Basel). 2014. PMID: 27135520 Free PMC article. Review.
-
A Rice CPYC-Type Glutaredoxin OsGRX20 in Protection against Bacterial Blight, Methyl Viologen and Salt Stresses.Front Plant Sci. 2018 Feb 9;9:111. doi: 10.3389/fpls.2018.00111. eCollection 2018. Front Plant Sci. 2018. PMID: 29479359 Free PMC article.
-
Comparative Transcriptomic Analysis Reveals That Ethylene/H2O2-Mediated Hypersensitive Response and Programmed Cell Death Determine the Compatible Interaction of Sand Pear and Alternaria alternata.Front Plant Sci. 2017 Feb 15;8:195. doi: 10.3389/fpls.2017.00195. eCollection 2017. Front Plant Sci. 2017. PMID: 28261248 Free PMC article.
-
Glutaredoxin AtGRXS8 represses transcriptional and developmental responses to nitrate in Arabidopsis thaliana roots.Plant Direct. 2020 Jun 11;4(6):e00227. doi: 10.1002/pld3.227. eCollection 2020 Jun. Plant Direct. 2020. PMID: 32537558 Free PMC article.
References
-
- Asada K. The water–water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annual Review of Plant Physiology and Plant Molecular Biology. 1999;50:601–639. - PubMed
-
- Asada K, Takahashi Y, Maeda Y, Enmanji K. Superoxide dismutases from a blue-green alga, Plectonema boryanum. Journal of Biological Chemistry. 1975;250:2801–2807. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
