

Tetrapyrrole synthesis of photosynthetic chromerids is likely homologous to the unusual pathway of apicomplexan parasites
- PMID: 21963666
- PMCID: PMC3203424
- DOI: 10.1105/tpc.111.089102
Tetrapyrrole synthesis of photosynthetic chromerids is likely homologous to the unusual pathway of apicomplexan parasites
Abstract
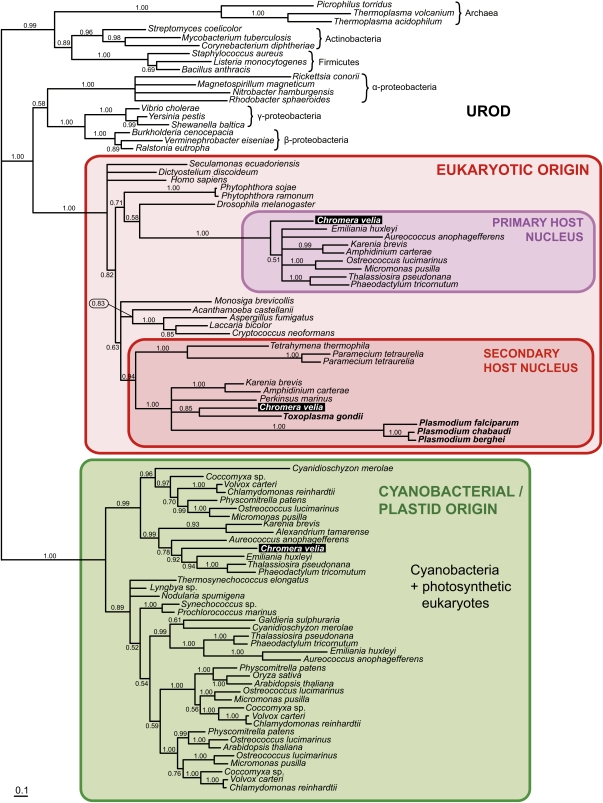
Most photosynthetic eukaryotes synthesize both heme and chlorophyll via a common tetrapyrrole biosynthetic pathway starting from glutamate. This pathway was derived mainly from cyanobacterial predecessor of the plastid and differs from the heme synthesis of the plastid-lacking eukaryotes. Here, we show that the coral-associated alveolate Chromera velia, the closest known photosynthetic relative to Apicomplexa, possesses a tetrapyrrole pathway that is homologous to the unusual pathway of apicomplexan parasites. We also demonstrate that, unlike other eukaryotic phototrophs, Chromera synthesizes chlorophyll from glycine and succinyl-CoA rather than glutamate. Our data shed light on the evolution of the heme biosynthesis in parasitic Apicomplexa and photosynthesis-related biochemical processes in their ancestors.
Figures



References
-
- Camadro J.M., Chambon H., Jolles J., Labbe P. (1986). Purification and properties of coproporphyrinogen oxidase from the yeast Saccharomyces cerevisiae. Eur. J. Biochem. 156: 579–587 - PubMed
-
- Emanuelsson O., Brunak S., von Heijne G., Nielsen H. (2007). Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2: 953–971 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
