

BAG-1 interacts with the p50-p50 homodimeric NF-κB complex: implications for colorectal carcinogenesis
- PMID: 21963853
- PMCID: PMC3272420
- DOI: 10.1038/onc.2011.452
BAG-1 interacts with the p50-p50 homodimeric NF-κB complex: implications for colorectal carcinogenesis
Abstract
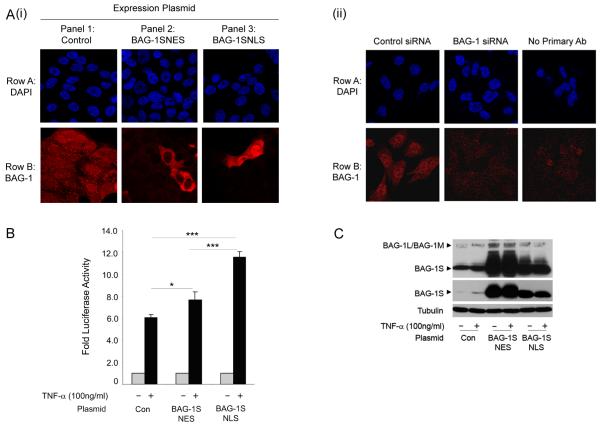
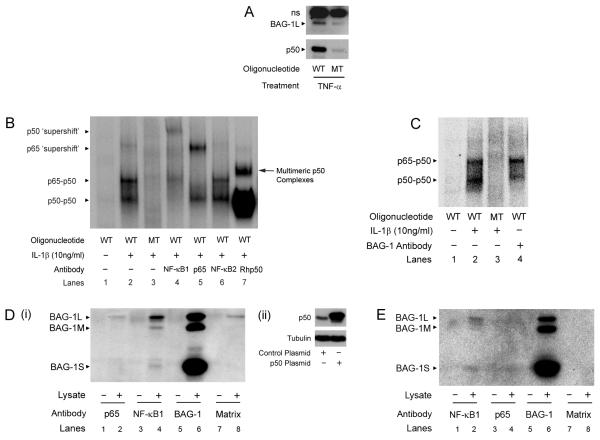
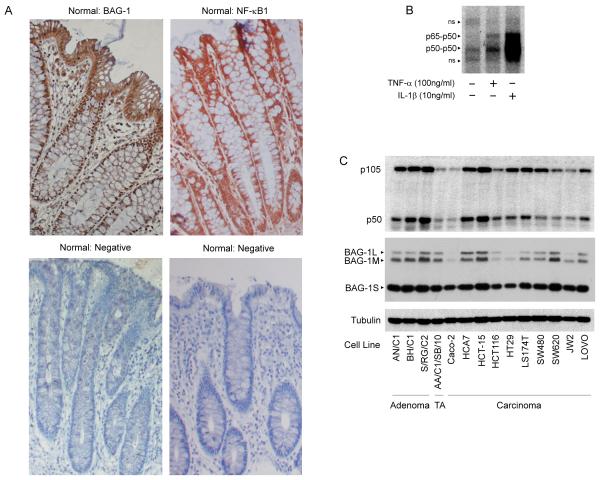
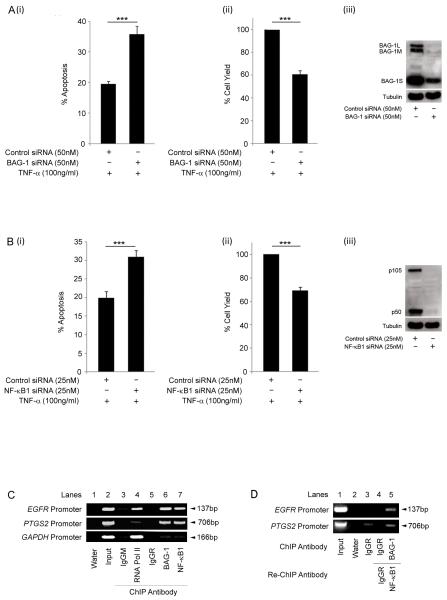
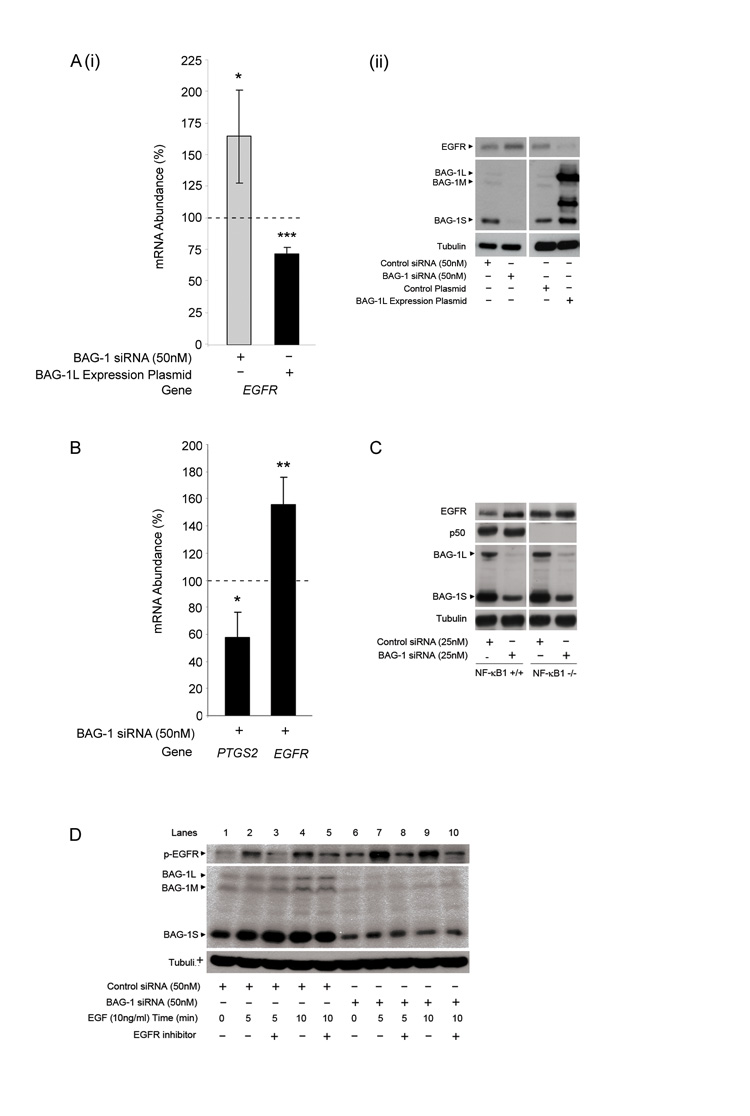
Understanding the mechanisms that promote aberrant tumour cell survival is critical for the determination of novel strategies to combat colorectal cancer (CRC). We have recently shown that the anti-apoptotic protein BAG-1, highly expressed in pre-malignant and CRC tissue, can potentiate cell survival through regulating NF-κB transcriptional activity. In this study, we identify a novel complex between BAG-1 and the p50-p50 NF-κB homodimers, implicating BAG-1 as a co-regulator of an atypical NF-κB pathway. Importantly, the BAG-1-p50 complex was detected at gene regulatory sequences including the epidermal growth factor receptor (EGFR) and COX-2 (PTGS2) genes. Suppression of BAG-1 expression using small interfering RNA was shown to increase EGFR and suppress COX-2 expression in CRC cells. Furthermore, mouse embryonic fibroblasts derived from the NF-κB1 (p105/p50) knock-out mouse were used to demonstrate that p50 expression was required for BAG-1 to suppress EGFR expression. This was shown to be functionally relevant as attenuation of BAG-1 expression increased ligand activated phosphorylation of EGFR in CRC cells. In summary, this paper identifies a novel role for BAG-1 in modulating gene expression through interaction with the p50-p50 NF-κB complexes. Data presented led us to propose that BAG-1 can act as a selective regulator of p50-p50 NF-κB responsive genes in colorectal tumour cells, potentially important for the promotion of cell survival in the context of the fluctuating tumour microenvironment. As BAG-1 expression is increased in the developing adenoma through to metastatic lesions, understanding the function of the BAG-1-p50 NF-κB complexes may aid in identifying strategies for both the prevention and treatment of CRC.
Figures
References
-
- Arhel NJ, Packham G, Townsend PA, Collard TJ, H-Zadeh AM, Sharp A, et al. The retinoblastoma protein interacts with BAG-1 in human colonic adenoma and carcinoma derived cell lines. Int J Cancer. 2003;106:364–71. - PubMed
-
- Arlt A, Vorndamm J, Muerkoster S, Yu H, Schmidt WE, Folsch UR, et al. Autocrine production of interleukin 1beta confers constitutive nuclear factor kappaB activity and chemoresistance in pancreatic carcinoma cell lines. Cancer Res. 2002;62:910–916. - PubMed
-
- Baeuerle PA, Baltimore D. I kappa B: a specific inhibitor of the NF-kappa B transcription factor. Science. 1988;242:540–546. - PubMed
-
- Barnes JD, Arhel NJ, Lee SS, Sharp A, Al-Okail M, Packham G, et al. Nuclear BAG-1 expression inhibits apoptosis in colorectal adenoma-derived epithelial cells. Apoptosis. 2005;10:301–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
