

Measuring mitochondrial function in intact cardiac myocytes
- PMID: 21964191
- PMCID: PMC3246130
- DOI: 10.1016/j.yjmcc.2011.08.030
Measuring mitochondrial function in intact cardiac myocytes
Abstract
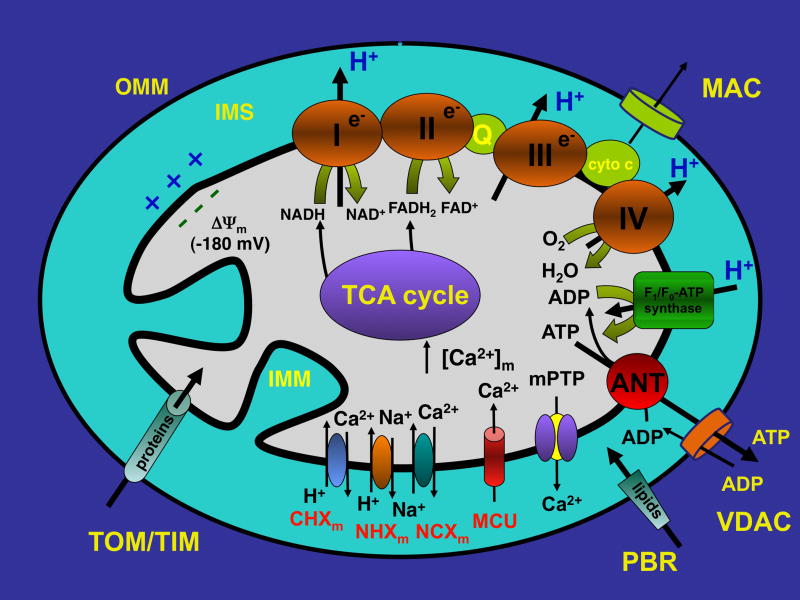
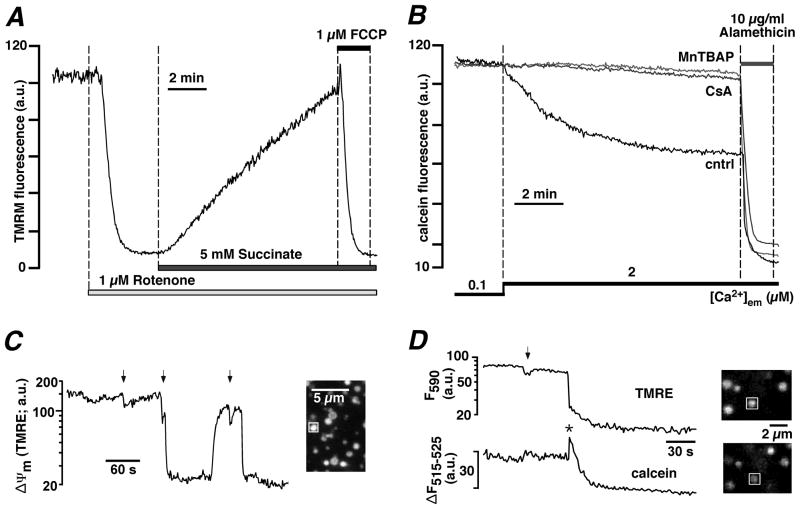
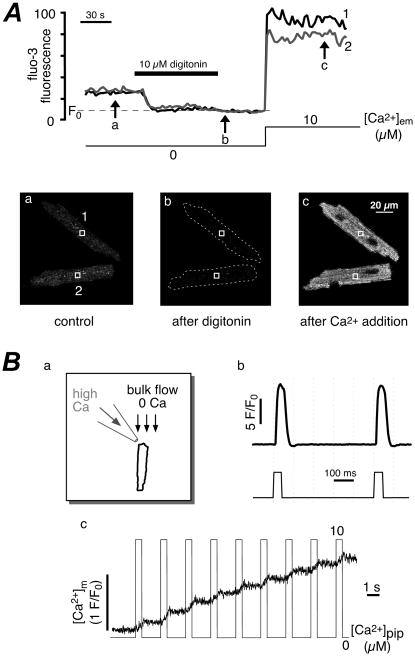
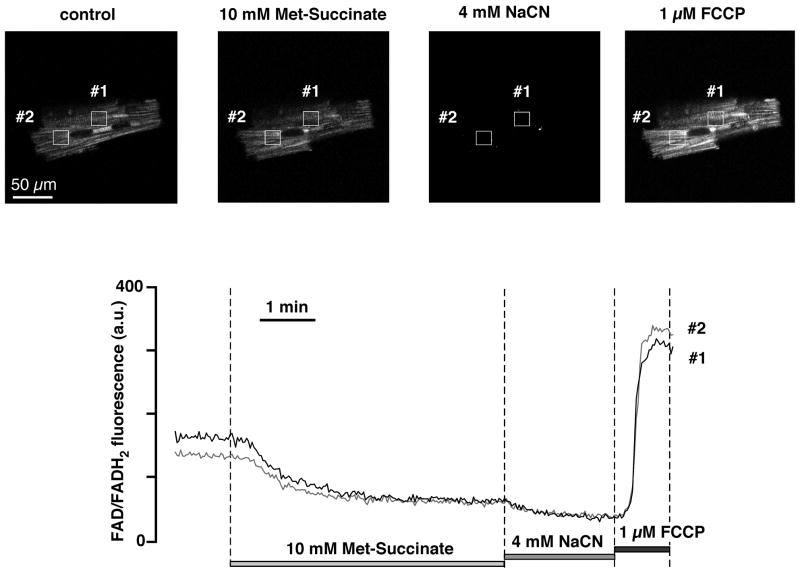
Mitochondria are involved in cellular functions that go beyond the traditional role of these organelles as the power plants of the cell. Mitochondria have been implicated in several human diseases, including cardiac dysfunction, and play a role in the aging process. Many aspects of our knowledge of mitochondria stem from studies performed on the isolated organelle. Their relative inaccessibility imposes experimental difficulties to study mitochondria in their natural environment-the cytosol of intact cells-and has hampered a comprehensive understanding of the plethora of mitochondrial functions. Here we review currently available methods to study mitochondrial function in intact cardiomyocytes. These methods primarily use different flavors of fluorescent dyes and genetically encoded fluorescent proteins in conjunction with high-resolution imaging techniques. We review methods to study mitochondrial morphology, mitochondrial membrane potential, Ca(2+) and Na(+) signaling, mitochondrial pH regulation, redox state and ROS production, NO signaling, oxygen consumption, ATP generation and the activity of the mitochondrial permeability transition pore. Where appropriate we complement this review on intact myocytes with seminal studies that were performed on isolated mitochondria, permeabilized cells, and in whole hearts.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
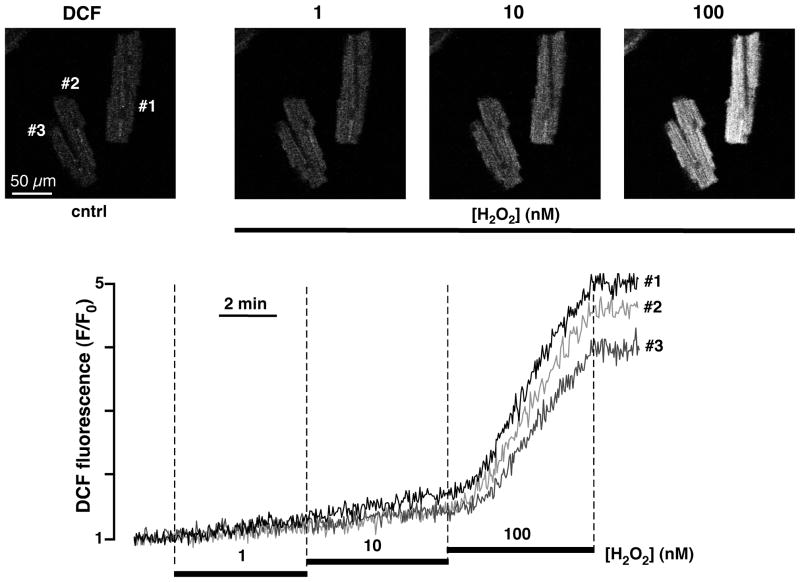
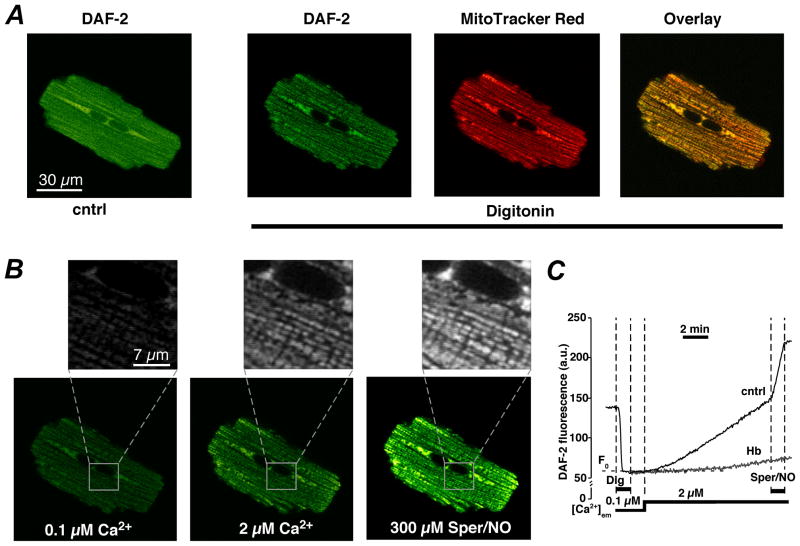
References
-
- Stanley WC, Recchia FA, Lopaschuk GD. Myocardial substrate metabolism in the normal and failing heart. Physiol Rev. 2005 Jul;85(3):1093–129. - PubMed
-
- McBride HM, Neuspiel M, Wasiak S. Mitochondria: more than just a powerhouse. Curr Biol. 2006 Jul 25;16(14):R551–60. - PubMed
-
- Barth E, Stammler G, Speiser B, Schaper J. Ultrastructural quantitation of mitochondria and myofilaments in cardiac muscle from 10 different animal species including man. J Mol Cell Cardiol. 1992 Jul;24(7):669–81. - PubMed
-
- Bers DM. Excitation-Contraction Coupling and Cardiac Contractile Force. 2. Dordrecht, Netherlands: Kluwer; 2001.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
