

Mucosal memory CD8⁺ T cells are selected in the periphery by an MHC class I molecule
- PMID: 21964609
- PMCID: PMC3197978
- DOI: 10.1038/ni.2106
Mucosal memory CD8⁺ T cells are selected in the periphery by an MHC class I molecule
Abstract
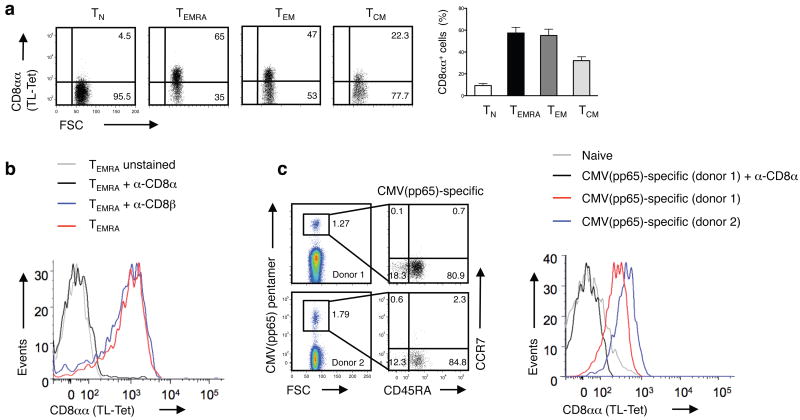
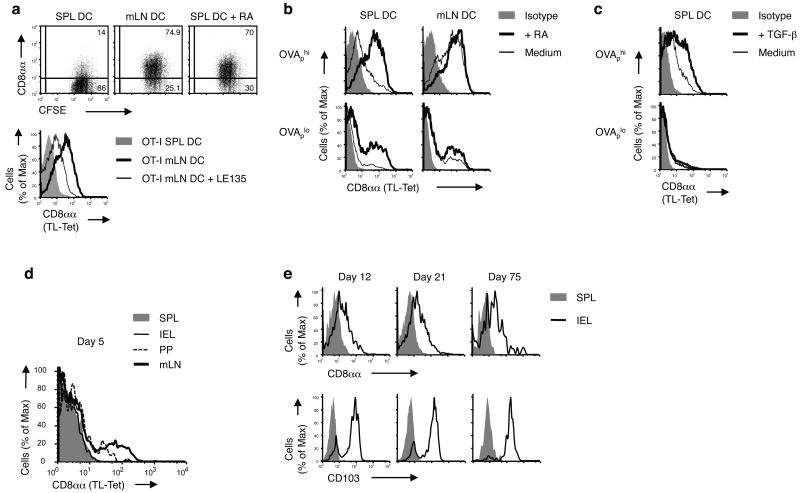
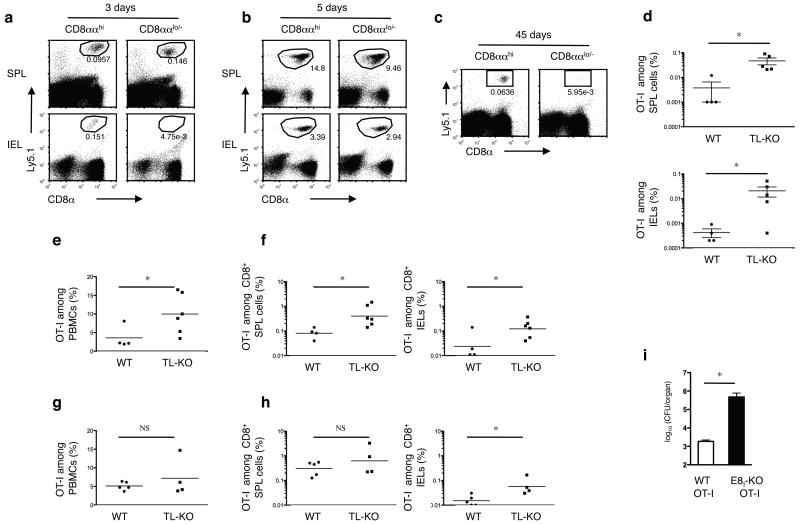
The presence of immune memory at pathogen-entry sites is a prerequisite for protection. Nevertheless, the mechanisms that warrant immunity at peripheral interfaces are not understood. Here we show that the nonclassical major histocompatibility complex (MHC) class I molecule thymus leukemia antigen (TL), induced on dendritic cells interacting with CD8αα on activated CD8αβ(+) T cells, mediated affinity-based selection of memory precursor cells. Furthermore, constitutive expression of TL on epithelial cells led to continued selection of mature CD8αβ(+) memory T cells. The memory process driven by TL and CD8αα was essential for the generation of CD8αβ(+) memory T cells in the intestine and the accumulation of highly antigen-sensitive CD8αβ(+) memory T cells that form the first line of defense at the largest entry port for pathogens.
Figures
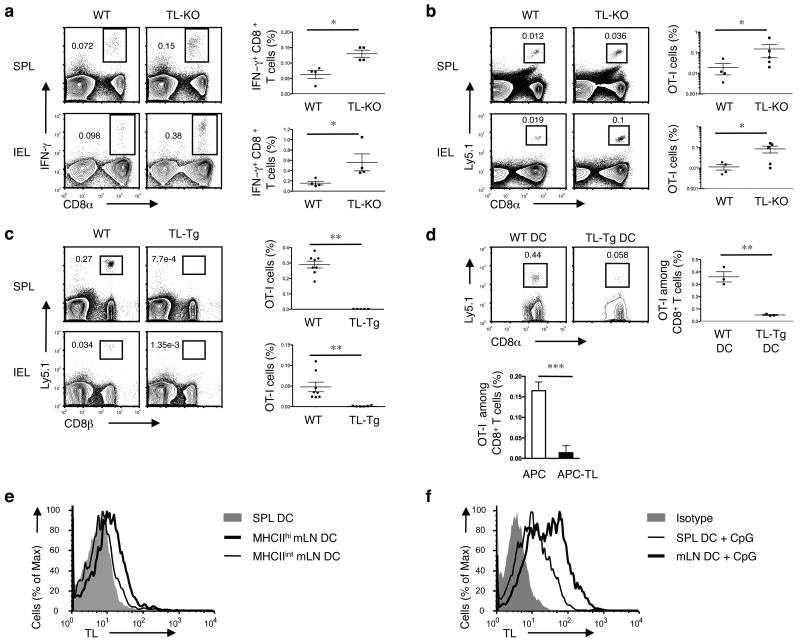
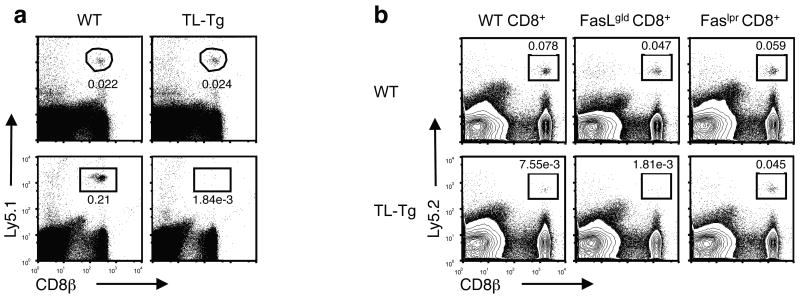
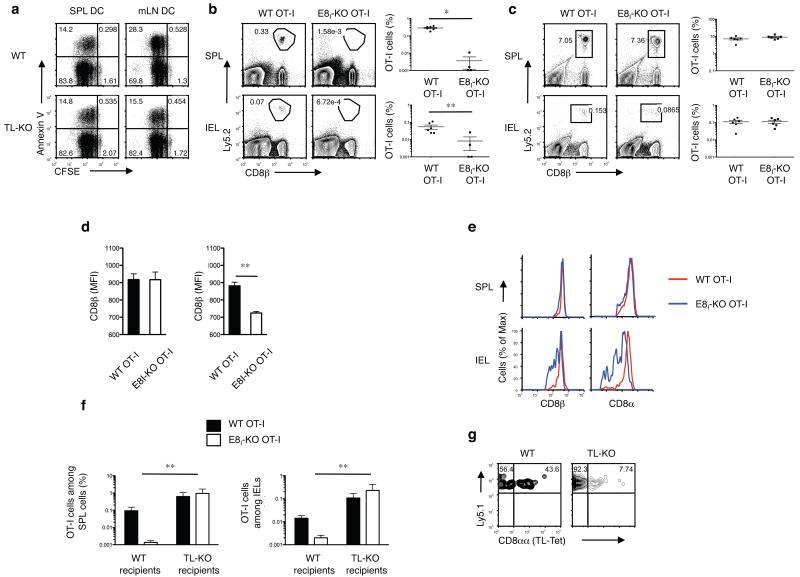
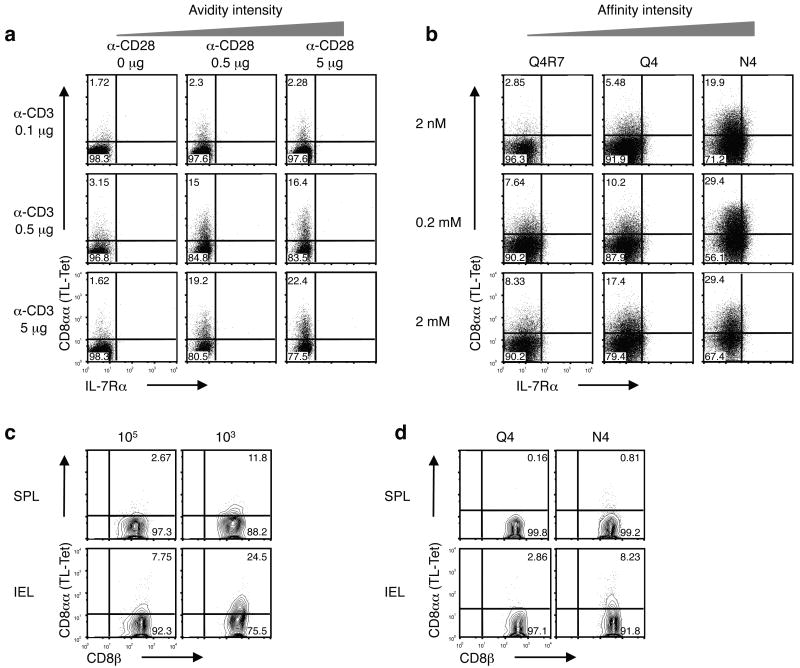
Comment in
-
TL we meet again.Nat Immunol. 2011 Oct 19;12(11):1027-8. doi: 10.1038/ni.2138. Nat Immunol. 2011. PMID: 22012434 No abstract available.
References
-
- Masopust D, Vezys V, Marzo AL, Lefrancois L. Preferential localization of effector memory cells in nonlymphoid tissue. Science. 2001;291:2413–2417. - PubMed
-
- Masopust D, Vezys V, Wherry EJ, Barber DL, Ahmed R. Cutting edge: gut microenvironment promotes differentiation of a unique memory CD8 T cell population. J Immunol. 2006;176:2079–2083. - PubMed
-
- Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
