

NMNAT suppresses tau-induced neurodegeneration by promoting clearance of hyperphosphorylated tau oligomers in a Drosophila model of tauopathy
- PMID: 21965302
- PMCID: PMC3276290
- DOI: 10.1093/hmg/ddr449
NMNAT suppresses tau-induced neurodegeneration by promoting clearance of hyperphosphorylated tau oligomers in a Drosophila model of tauopathy
Abstract
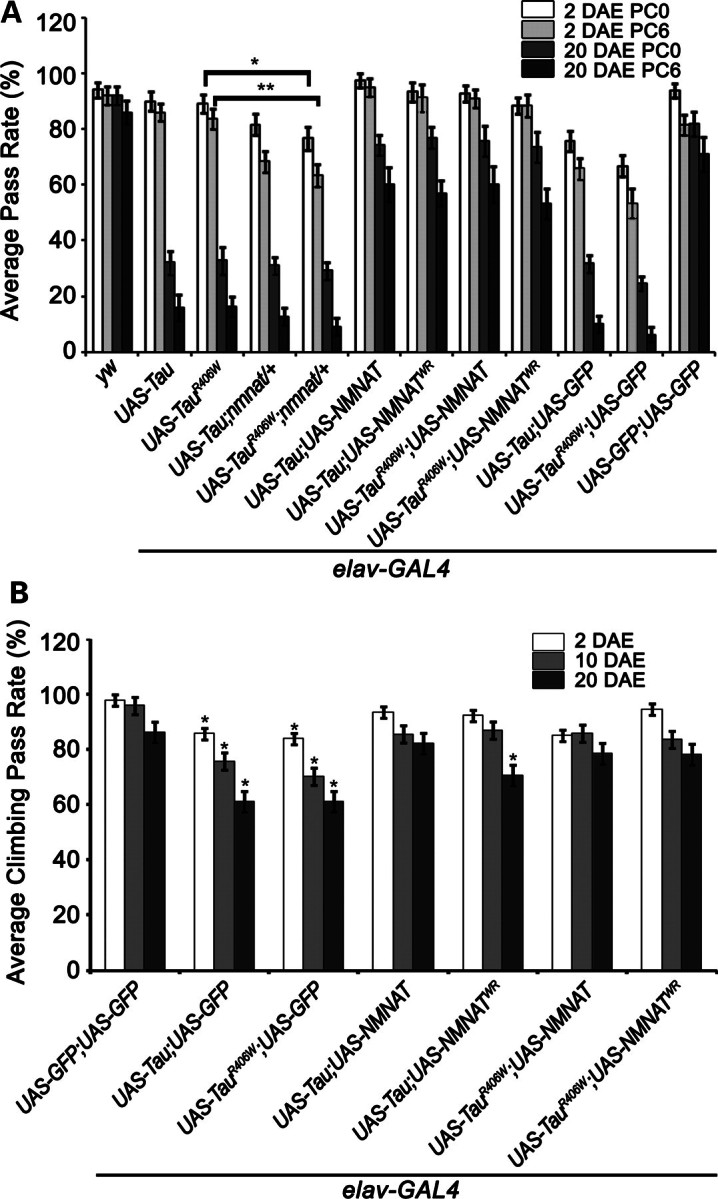
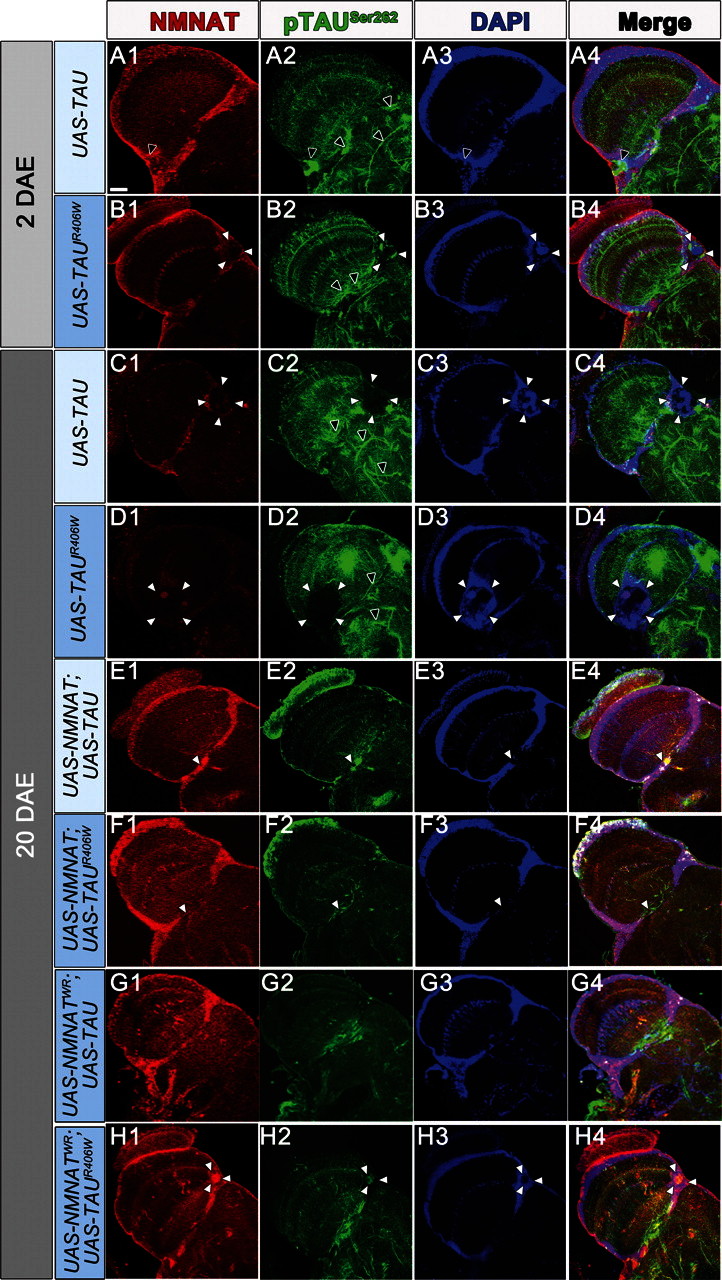
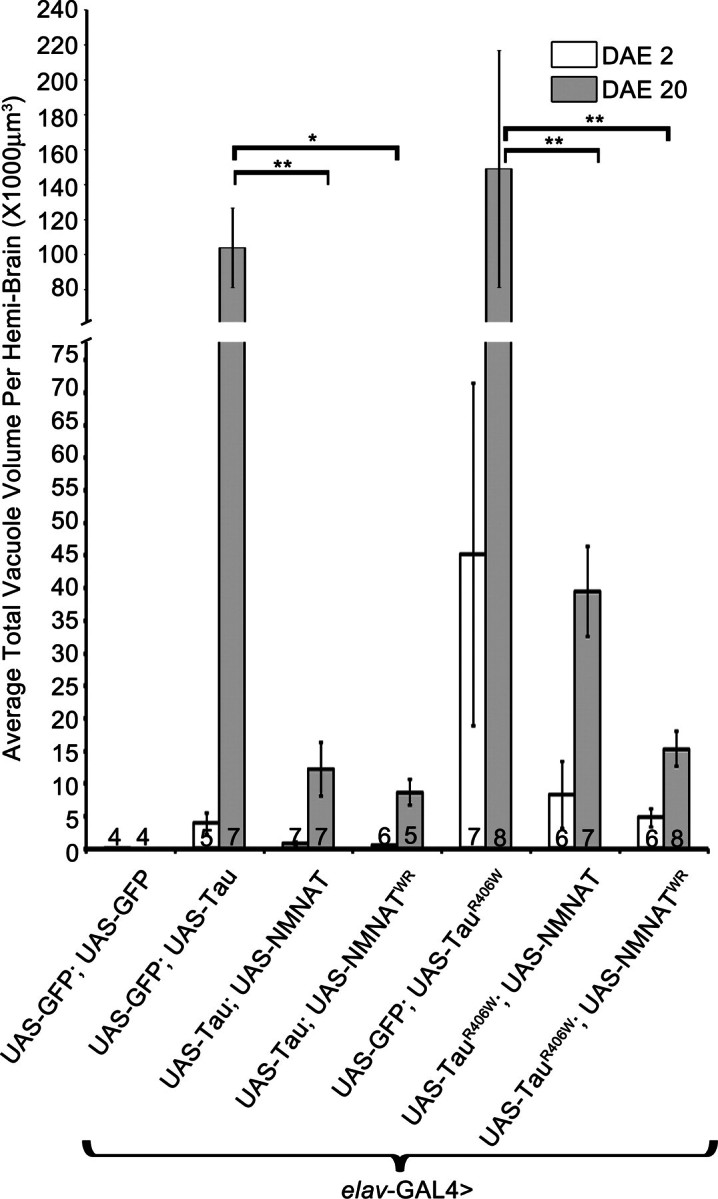
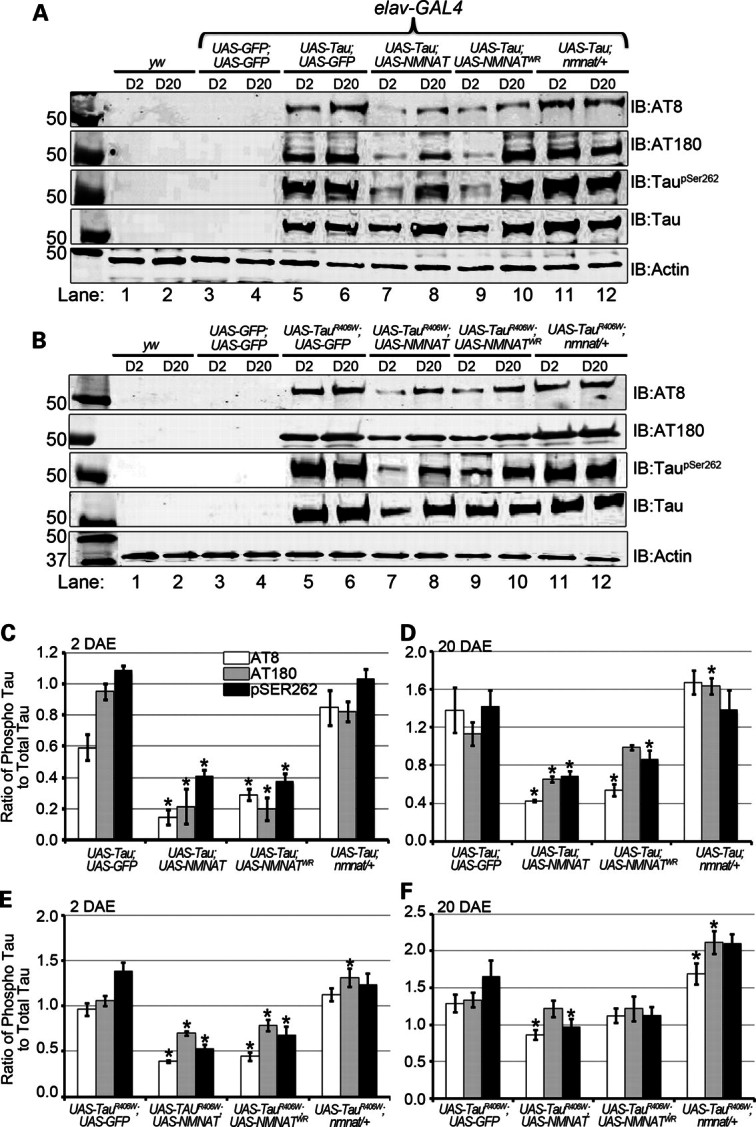
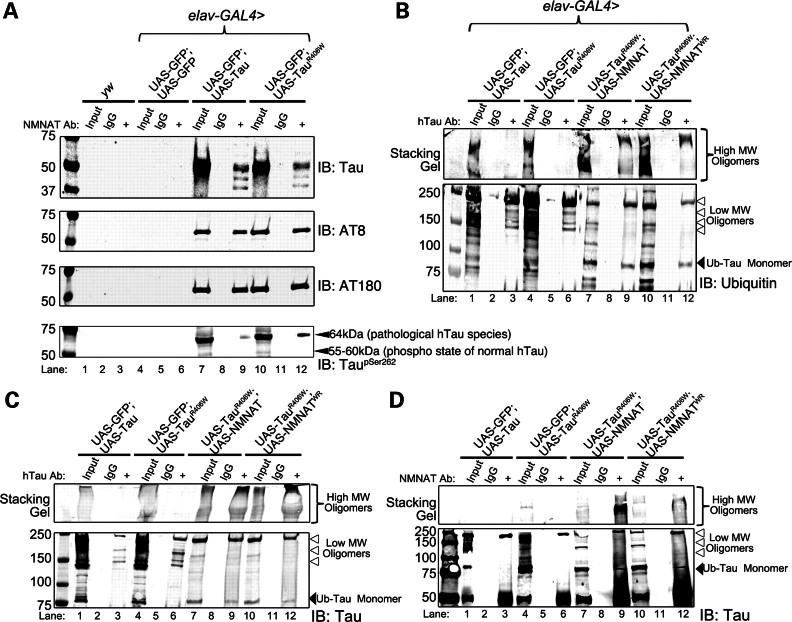
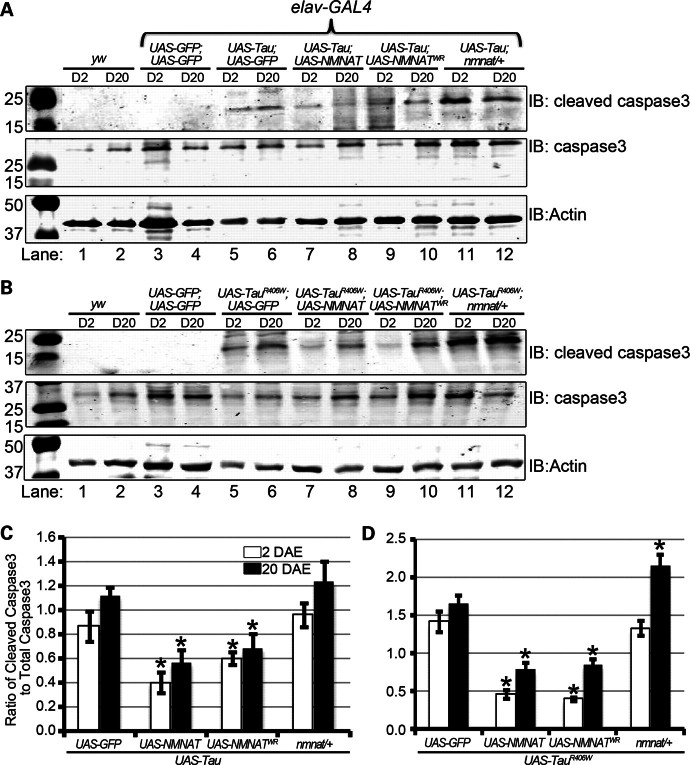
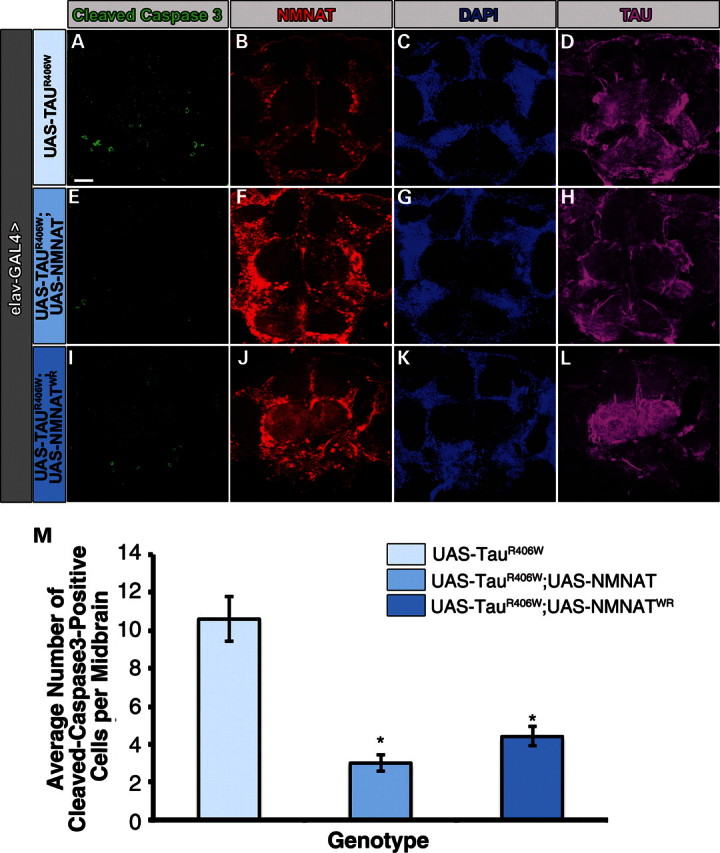
Tauopathies, including Alzheimer's disease, are a group of neurodegenerative diseases characterized by abnormal tau hyperphosphorylation that leads to formation of neurofibrillary tangles. Drosophila models of tauopathy display prominent features of the human disease including compromised lifespan, impairments of learning, memory and locomotor functions and age-dependent neurodegeneration visible as vacuolization. Here, we use a Drosophila model of frontotemporal dementia with parkinsonism linked to chromosome 17 (FTDP-17), in order to study the neuroprotective capacity of a recently identified neuronal maintenance factor, nicotinamide mononucleotide (NAD) adenylyl transferase (NMNAT), a protein that has both NAD synthase and chaperone function. NMNAT is essential for maintaining neuronal integrity under normal conditions and has been shown to protect against several neurodegenerative conditions. However, its protective role in tauopathy has not been examined. Here, we show that overexpression of NMNAT significantly suppresses both behavioral and morphological deficits associated with tauopathy by means of reducing the levels of hyperphosphorylated tau oligomers. Importantly, the protective activity of NMNAT protein is independent of its NAD synthesis activity, indicating a role for direct protein-protein interaction. Next, we show that NMNAT interacts with phosphorylated tau in vivo and promotes the ubiquitination and clearance of toxic tau species. Consequently, apoptosis activation was significantly reduced in brains overexpressing NMNAT, and neurodegeneration was suppressed. Our report on the molecular basis of NMNAT-mediated neuroprotection in tauopathies opens future investigation of this factor in other protein foldopathies.
© The Author 2011. Published by Oxford University Press. All rights reserved.
Figures
References
-
- Iqbal K., Alonso Adel C., Chen S., Chohan M.O., El-Akkad E., Gong C.X., Khatoon S., Li B., Liu F., Rahman A., et al. Tau pathology in Alzheimer disease and other tauopathies. Biochim. Biophys. Acta. 2005;1739:198–210. - PubMed
-
- Sergeant N., Delacourte A., Buee L. Tau protein as a differential biomarker of tauopathies. Biochim. Biophys. Acta. 2005;1739:179–197. - PubMed
-
- Alonso A.C., Grundke-Iqbal I., Iqbal K. Alzheimer's disease hyperphosphorylated tau sequesters normal tau into tangles of filaments and disassembles microtubules. Nat. Med. 1996;2:783–787. - PubMed
-
- Khatoon S., Grundke-Iqbal I., Iqbal K. Brain levels of microtubule-associated protein tau are elevated in Alzheimer's disease: a radioimmuno-slot-blot assay for nanograms of the protein. J. Neurochem. 1992;59:750–753. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
