

Saccharomyces cerevisiae Ngl3p is an active 3'-5' exonuclease with a specificity towards poly-A RNA reminiscent of cellular deadenylases
- PMID: 21965533
- PMCID: PMC3258157
- DOI: 10.1093/nar/gkr782
Saccharomyces cerevisiae Ngl3p is an active 3'-5' exonuclease with a specificity towards poly-A RNA reminiscent of cellular deadenylases
Abstract
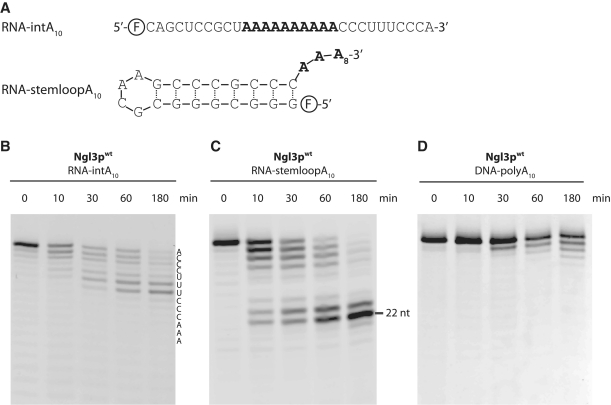
Deadenylation is the first and rate-limiting step during turnover of mRNAs in eukaryotes. In the yeast, Saccharomyces cerevisiae, two distinct 3'-5' exonucleases, Pop2p and Ccr4p, have been identified within the Ccr4-NOT deadenylase complex, belonging to the DEDD and Exonuclease-Endonuclease-Phosphatase (EEP) families, respectively. Ngl3p has been identified as a new member of the EEP family of exonucleases based on sequence homology, but its activity and biological roles are presently unknown. Here, we show using in vitro deadenylation assays on defined RNA species mimicking poly-A containing mRNAs that yeast Ngl3p is a functional 3'-5' exonuclease most active at slightly acidic conditions. We further show that the enzyme depends on divalent metal ions for activity and possesses specificity towards poly-A RNA similar to what has been observed for cellular deadenylases. The results suggest that Ngl3p is naturally involved in processing of poly-adenylated RNA and provide insights into the mechanistic variations observed among the redundant set of EEP enzymes found in yeast and higher eukaryotes.
Figures




References
-
- Goldstrohm AC, Wickens M. Multifunctional deadenylase complexes diversify mRNA control. Nat. Rev. Mol. Cell Biol. 2008;9:337–344. - PubMed
-
- Parker R, Song H. The enzymes and control of eukaryotic mRNA turnover. Nat. Struct. Mol. Biol. 2004;11:121–127. - PubMed
-
- Houseley J, Tollervey D. The many pathways of RNA degradation. Cell. 2009;136:763–776. - PubMed
-
- Garneau NL, Wilusz J, Wilusz CJ. The highways and byways of mRNA decay. Nat. Rev. Mol. Cell Biol. 2007;8:113–126. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
