

Regulation of biofilm components in Salmonella enterica serovar Typhimurium by lytic transglycosylases involved in cell wall turnover
- PMID: 21965572
- PMCID: PMC3232906
- DOI: 10.1128/JB.00425-11
Regulation of biofilm components in Salmonella enterica serovar Typhimurium by lytic transglycosylases involved in cell wall turnover
Abstract
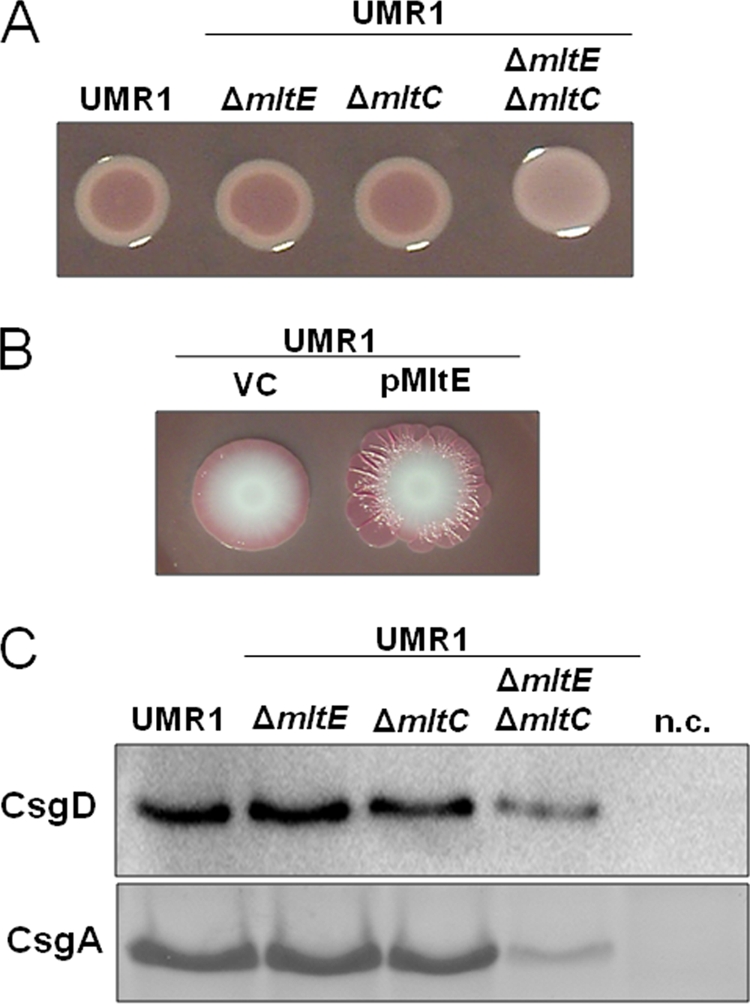
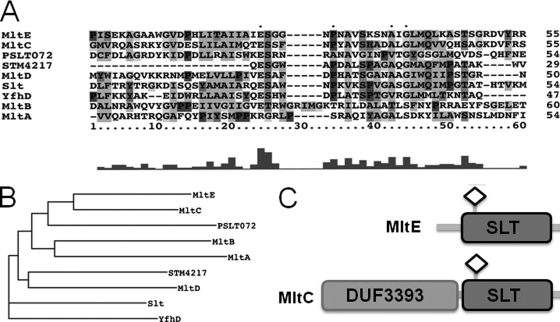
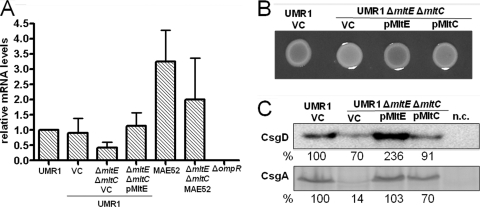
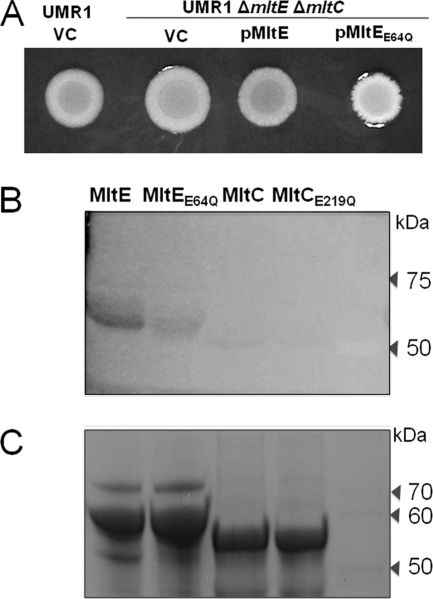
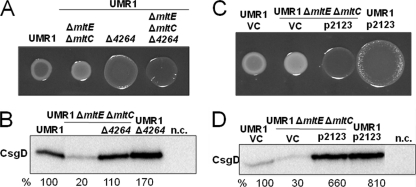
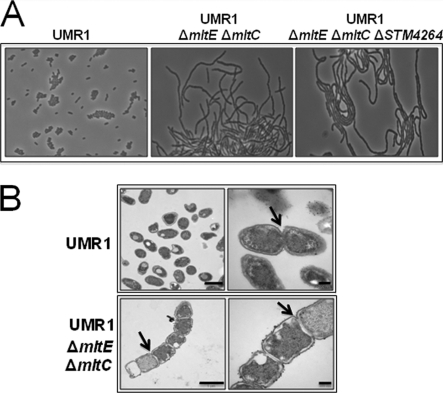
In Salmonella enterica serovar Typhimurium, a biofilm mode of growth known as the rdar morphotype is regulated by several networks which sense multiple environmental signals. The transcriptional regulator CsgD is the major target for these regulatory pathways. In this study, we show that two lytic transglycosylases of family I, MltE and MltC, in combination increase CsgD expression and rdar morphotype. MltE and MltC, which share a highly similar transglycosylase SLT domain, work redundantly to regulate CsgD at the transcriptional and posttranscriptional levels. The effect of MltE and MltC on CsgD levels was independent of the known regulatory pathways that sense cell envelope stress. These findings reveal, for the first time, a specific function of lytic transglycosylases in S. Typhimurium and suggest the existence of a new signaling pathway that links cell wall turnover to biofilm formation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
