

Members of the salivary gland surface protein (SGS) family are major immunogenic components of mosquito saliva
- PMID: 21965675
- PMCID: PMC3220476
- DOI: 10.1074/jbc.M111.280552
Members of the salivary gland surface protein (SGS) family are major immunogenic components of mosquito saliva
Abstract
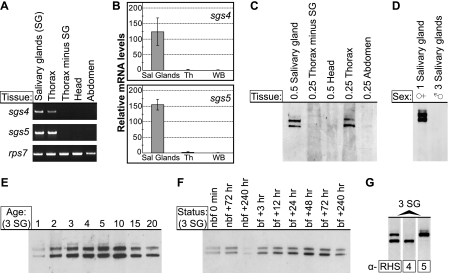
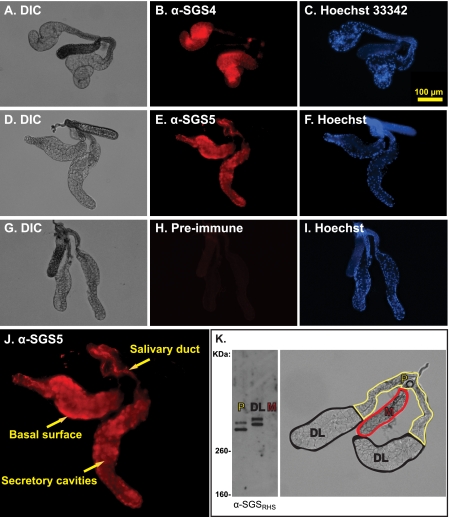
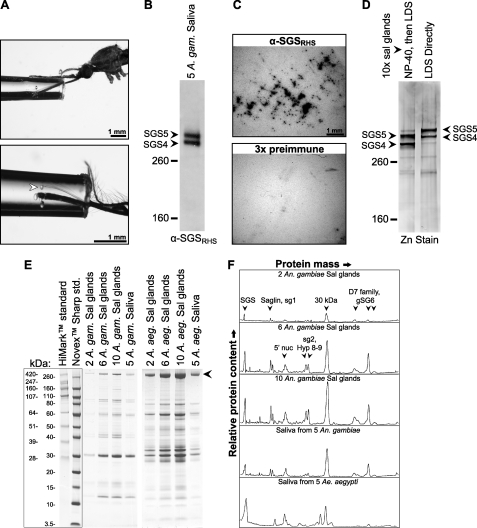
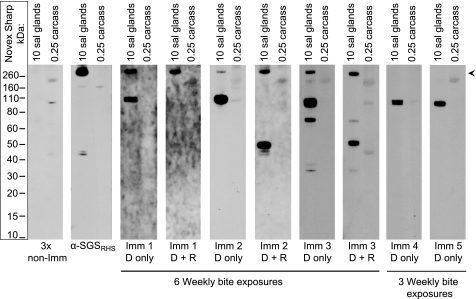
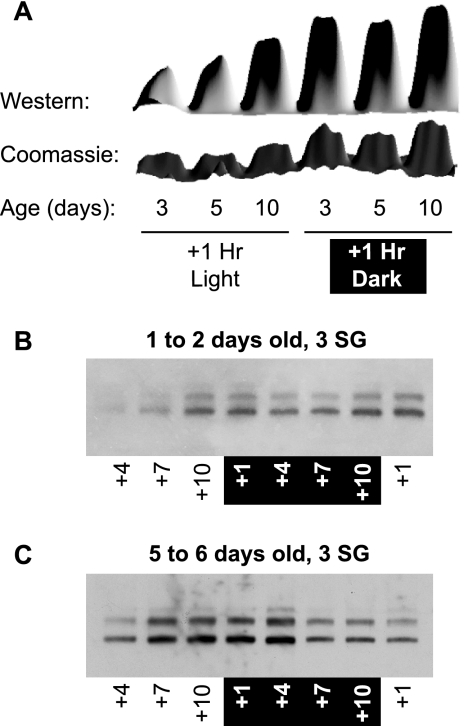
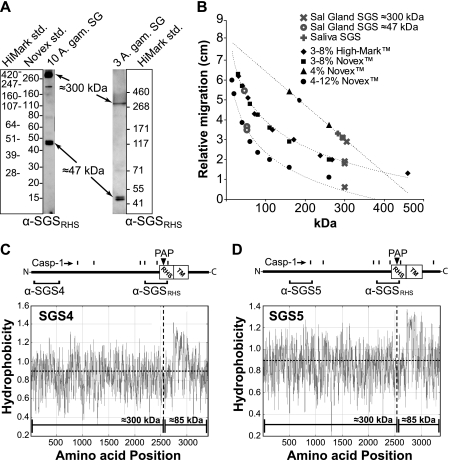
Mosquitoes transmit Plasmodium and certain arboviruses during blood feeding, when they are injected along with saliva. Mosquito saliva interferes with the host's hemostasis and inflammation response and influences the transmission success of some pathogens. One family of mosquito salivary gland proteins, named SGS, is composed of large bacterial-type proteins that in Aedes aegypti were implicated as receptors for Plasmodium on the basal salivary gland surface. Here, we characterize the biology of two SGSs in the malaria mosquito, Anopheles gambiae, and demonstrate their involvement in blood feeding. Western blots and RT-PCR showed that Sgs4 and Sgs5 are produced exclusively in female salivary glands, that expression increases with age and after blood feeding, and that protein levels fluctuate in a circadian manner. Immunohistochemistry showed that SGSs are present in the acinar cells of the distal lateral lobes and in the salivary ducts of the proximal lobes. SDS-PAGE, Western blots, bite blots, and immunization via mosquito bites showed that SGSs are highly immunogenic and form major components of mosquito saliva. Last, Western and bioinformatic analyses suggest that SGSs are secreted via a non-classical pathway that involves cleavage into a 300-kDa soluble fragment and a smaller membrane-bound fragment. Combined, these data strongly suggest that SGSs play an important role in blood feeding. Together with their role in malaria transmission, we propose that SGSs could be used as markers of human exposure to mosquito bites and in the development of disease control strategies.
Figures
Similar articles
-
Natural human humoral response to salivary gland proteins of Anopheles mosquitoes in Thailand.Acta Trop. 2006 Apr;98(1):66-73. doi: 10.1016/j.actatropica.2006.02.004. Epub 2006 Mar 10. Acta Trop. 2006. PMID: 16530153
-
Is Saglin a mosquito salivary gland receptor for Plasmodium falciparum?Malar J. 2019 Jan 3;18(1):2. doi: 10.1186/s12936-018-2634-5. Malar J. 2019. PMID: 30602380 Free PMC article.
-
Salivary gland-specific gene expression in the malaria vector Anopheles gambiae.Parassitologia. 1999 Sep;41(1-3):483-7. Parassitologia. 1999. PMID: 10697906 Review.
-
A mosquito-specific protein family includes candidate receptors for malaria sporozoite invasion of salivary glands.Cell Microbiol. 2006 Jan;8(1):163-75. doi: 10.1111/j.1462-5822.2005.00611.x. Cell Microbiol. 2006. PMID: 16367875
-
Insect-malaria parasites interactions: the salivary gland.Insect Biochem Mol Biol. 2004 Jul;34(7):615-24. doi: 10.1016/j.ibmb.2004.03.014. Insect Biochem Mol Biol. 2004. PMID: 15242702 Review.
Cited by
-
Identification of Aedes aegypti salivary gland proteins interacting with human immune receptor proteins.PLoS Negl Trop Dis. 2022 Sep 7;16(9):e0010743. doi: 10.1371/journal.pntd.0010743. eCollection 2022 Sep. PLoS Negl Trop Dis. 2022. PMID: 36070318 Free PMC article.
-
Serological responses and biomarker evaluation in mice and pigs exposed to tsetse fly bites.PLoS Negl Trop Dis. 2014 May 22;8(5):e2911. doi: 10.1371/journal.pntd.0002911. eCollection 2014 May. PLoS Negl Trop Dis. 2014. PMID: 24853371 Free PMC article.
-
Plasmodium berghei sporozoites acquire virulence and immunogenicity during mosquito hemocoel transit.Infect Immun. 2014 Mar;82(3):1164-72. doi: 10.1128/IAI.00758-13. Epub 2013 Dec 30. Infect Immun. 2014. PMID: 24379288 Free PMC article.
-
Genetic changes of Plasmodium vivax tempers host tissue-specific responses in Anopheles stephensi.Curr Res Immunol. 2021 Feb 20;2:12-22. doi: 10.1016/j.crimmu.2021.02.002. eCollection 2021. Curr Res Immunol. 2021. PMID: 35492403 Free PMC article.
-
Wolbachia variants induce differential protection to viruses in Drosophila melanogaster: a phenotypic and phylogenomic analysis.PLoS Genet. 2013;9(12):e1003896. doi: 10.1371/journal.pgen.1003896. Epub 2013 Dec 12. PLoS Genet. 2013. PMID: 24348259 Free PMC article.
References
-
- Mans B. J., Francischetti I. M. (2011) in Toxins and Hemostasis (Kini R. M., Clemetson K. J., Markland F. S., McLane M. A., Morita T. eds) pp. 21–44, Springer Science + Business Media, New York
-
- Ribeiro J. M. C., Arcà B. (2009) Adv. Insect Physiol. 37, 59–118
-
- Remoue F., Cisse B., Ba F., Sokhna C., Herve J. P., Boulanger D., Simondon F. (2006) Trans. R. Soc. Trop. Med. Hyg. 100, 363–370 - PubMed
-
- Titus R. G., Bishop J. V., Mejia J. S. (2006) Parasite Immunol. 28, 131–141 - PubMed
-
- World Health Organization (1996) Weekly Epidemiological Record 71, 17–22 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
