

Actions of Tefluthrin on Rat Na(v)1.7 Voltage-Gated Sodium Channels Expressed in Xenopus Oocytes
- PMID: 21966053
- PMCID: PMC3181098
- DOI: 10.1016/j.pestbp.2011.06.001
Actions of Tefluthrin on Rat Na(v)1.7 Voltage-Gated Sodium Channels Expressed in Xenopus Oocytes
Abstract
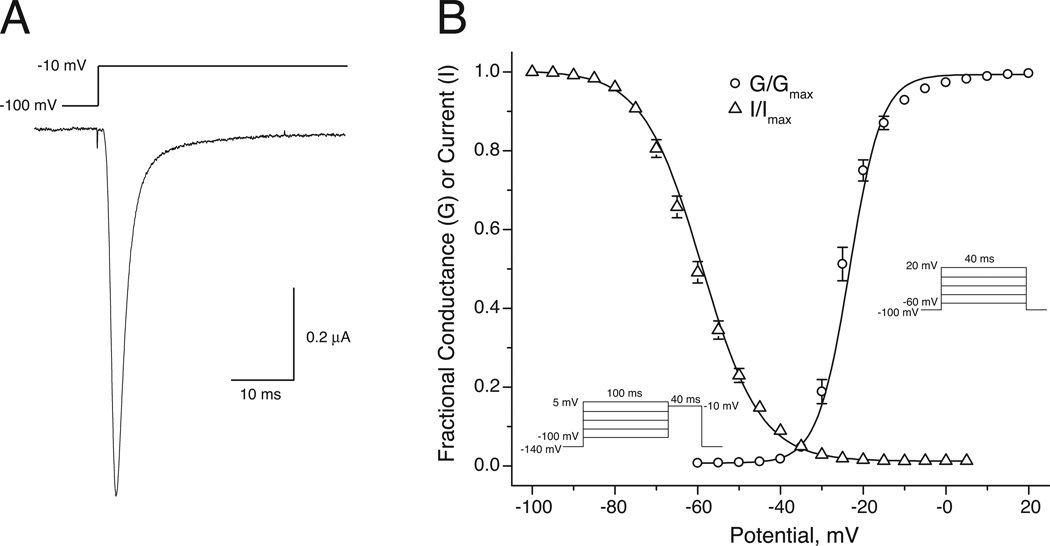
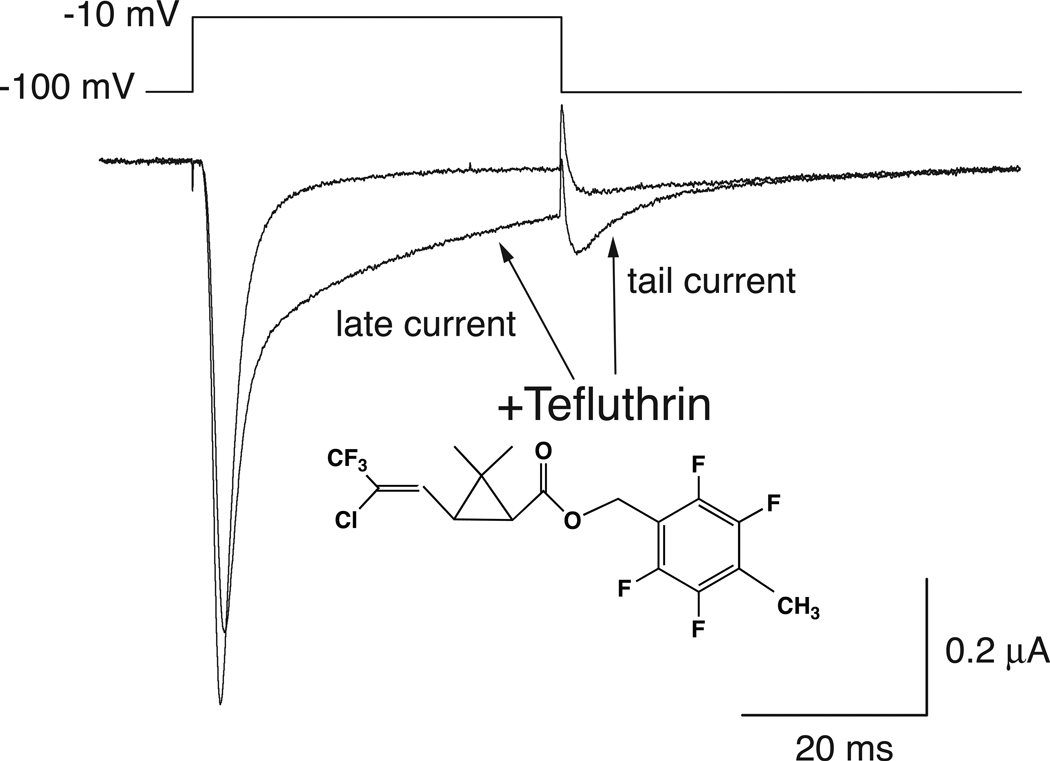
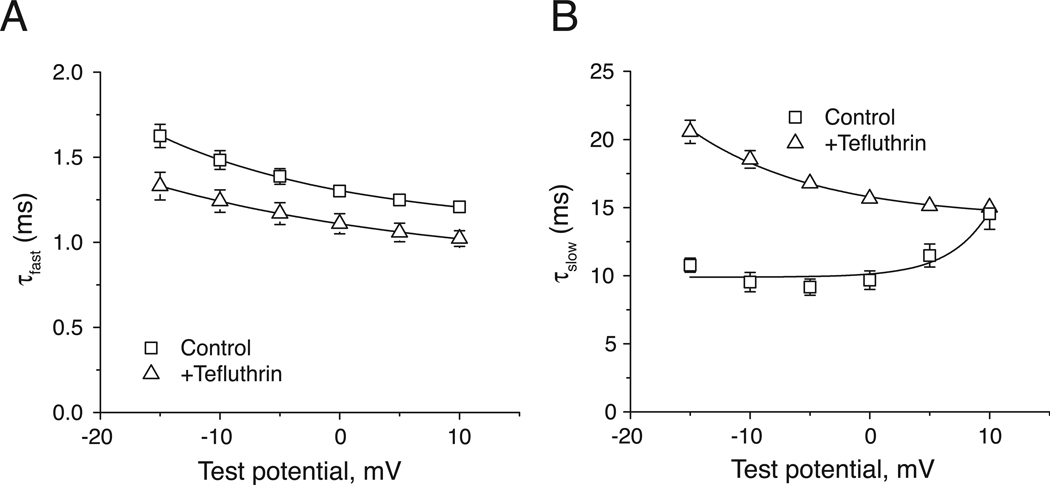
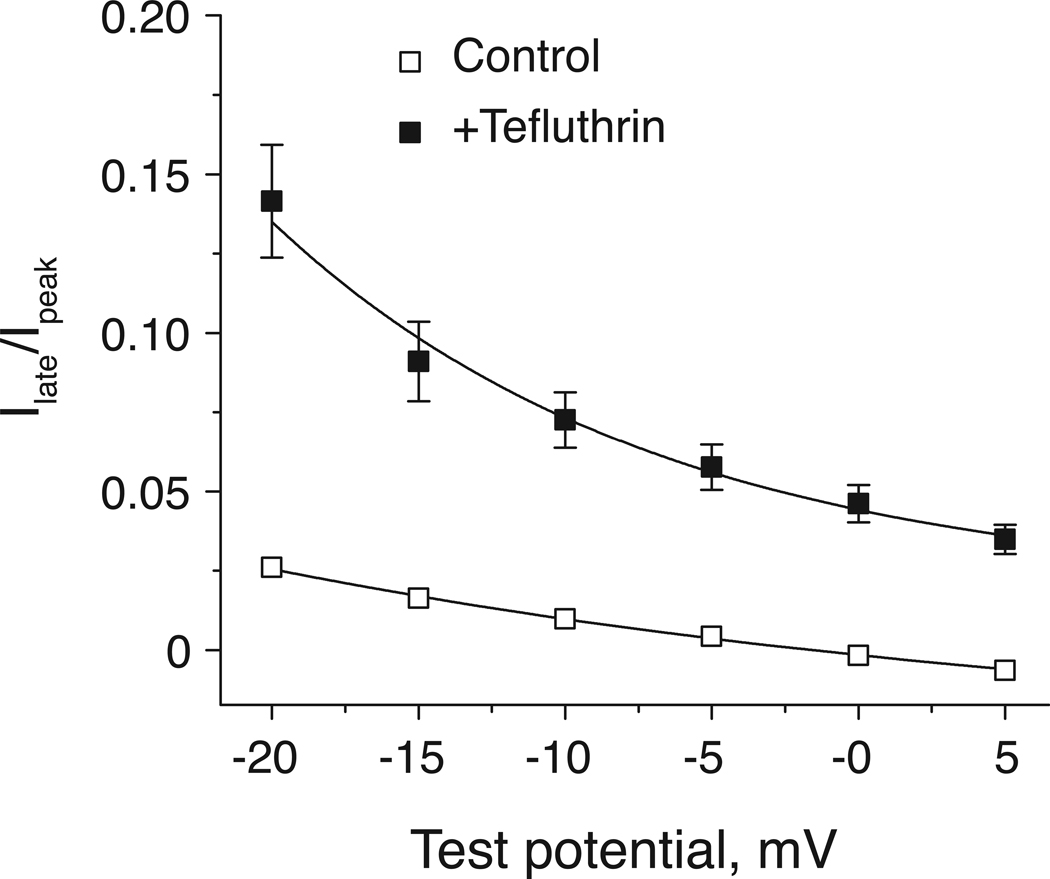
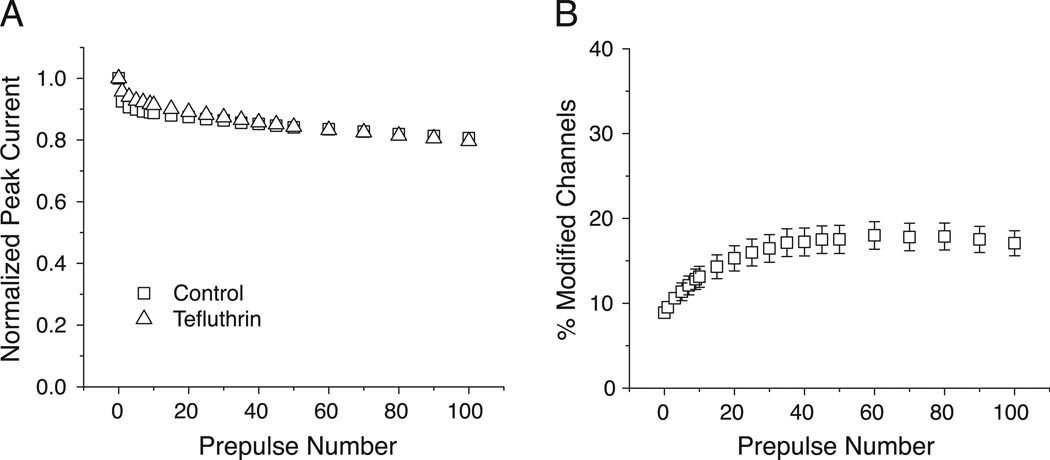
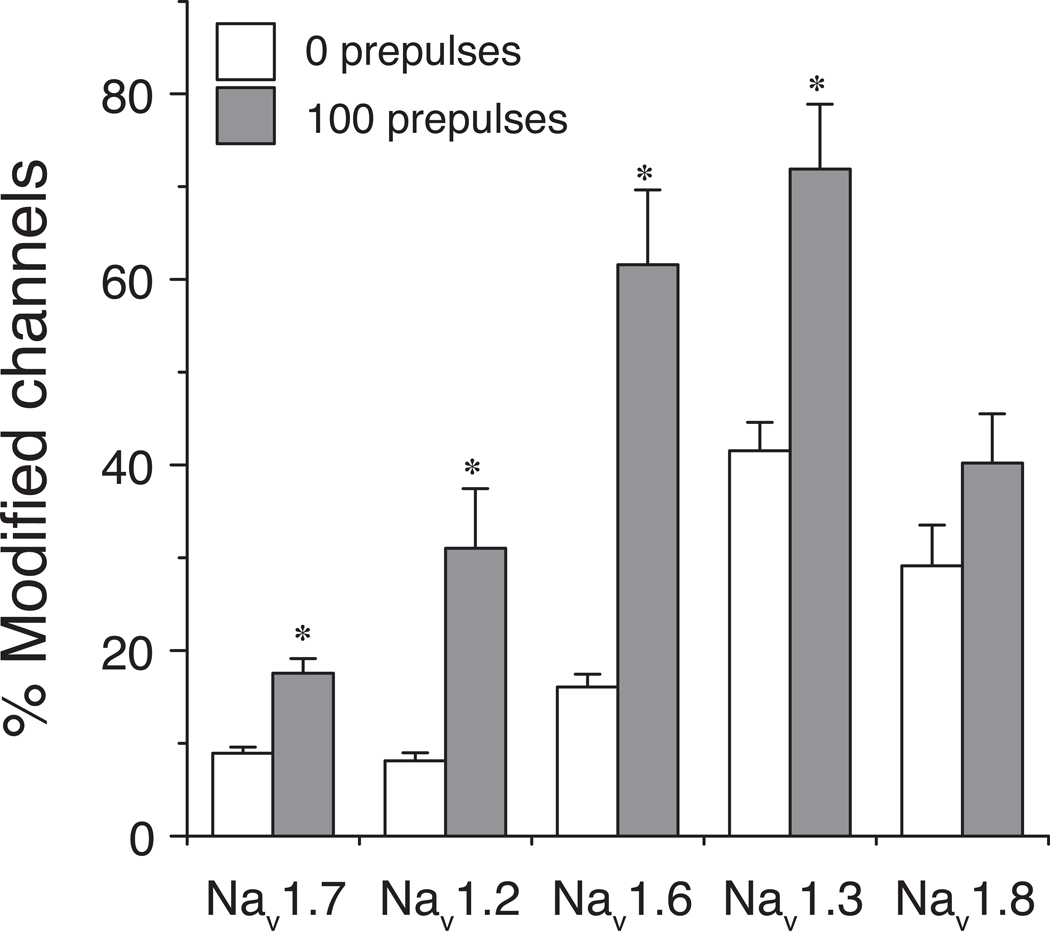
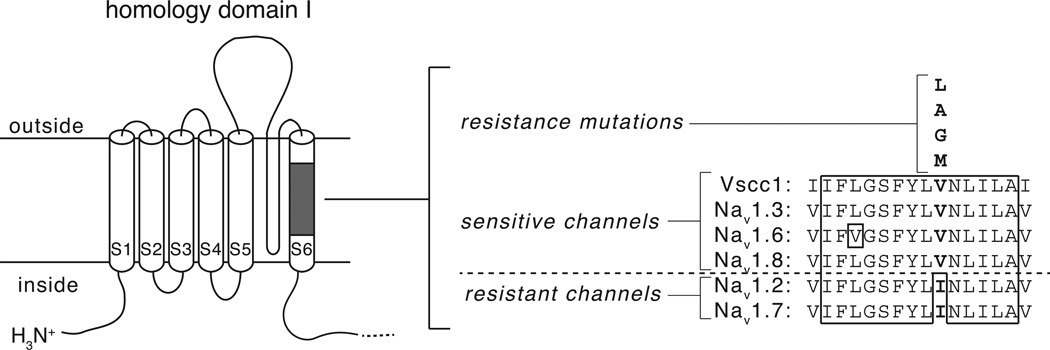
In rats expression of the Na(v)1.7 voltage-gated sodium channel isoform is restricted to the peripheral nervous system and is abundant in the sensory neurons of the dorsal root ganglion. We expressed the rat Na(v)1.7 sodium channel α subunit together with the rat auxiliary β1 and β2 subunits in Xenopus laevis oocytes and assessed the effects of the pyrethroid insecticide tefluthrin on the expressed currents using the two-electrode voltage clamp method. Tefluthrin at 100 µM modified of Na(v)1.7 channels to prolong inactivation of the peak current during a depolarizing pulse, resulting in a marked "late current" at the end of a 40-ms depolarization, and induced a sodium tail current following repolarization. Tefluthrin modification was enhanced up to two-fold by the application of a train of up to 100 5-ms depolarizing prepulses. These effects of tefluthrin on Na(v)1.7 channels were qualitatively similar to its effects on rat Na(v)1.2, Na(v)1.3 and Na(v)1.6 channels assayed previously under identical conditions. However, Na(v)1.7 sodium channels were distinguished by their low sensitivity to modification by tefluthrin, especially compared to Na(v)1.3 and Na(v)1.6 channels. It is likely that Na(v)1.7 channels contribute significantly to the tetrodotoxin-sensitive, pyrethroid-resistant current found in cultured dorsal root ganglion neurons. We aligned the complete amino acid sequences of four pyrethroid-sensitive isoforms (house fly Vssc1; rat Na(v)1.3, Na(v)1.6 and Na(v)1.8) and two pyrethroid-resistant isoforms (rat Na(v)1.2 and Na(v)1.7) and found only a single site, located in transmembrane segment 6 of homology domain I, at which the amino acid sequence was conserved among all four sensitive isoform sequences but differed in the two resistant isoform sequences. This position, corresponding to Val410 of the house fly Vssc1 sequence, also aligns with sites of multiple amino acid substitutions identified in the sodium channel sequences of pyrethroid-resistant insect populations. These results implicate this single amino acid polymorphism in transmembrane segment 6 of sodium channel homology domain I as a determinant of the differential pyrethroid sensitivity of rat sodium channel isoforms.
Figures
References
-
- Soderlund DM, Clark JM, Sheets LP, Mullin LS, Piccirillo VJ, Sargent D, Stevens JT, Weiner ML. Mechanisms of pyrethroid toxicity: implications for cumulative risk assessment. Toxicology. 2002;171:3–59. - PubMed
-
- Soderlund DM. Mode of action of pyrethrins and pyrethroids. In: Casida JE, Quistad GB, editors. Pyrethrum Flowers: Production, Chemistry, Toxicology, and Uses. New York: Oxford University Press; 1995. pp. 217–233.
-
- Catterall WA. From ionic currents to molecular mechanisms: structure and function of voltage-gated sodium channels. Neuron. 2000;26:13–25. - PubMed
-
- Goldin AL. Resurgence of sodium channel research. Annual Review of Physiology. 2001;63:871–894. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources