

Review
doi: 10.1083/jcb.201103148.
Pathogens and polymers: microbe-host interactions illuminate the cytoskeleton
Affiliations
- PMID: 21969466
- PMCID: PMC3187711
- DOI: 10.1083/jcb.201103148
Item in Clipboard
Review
Pathogens and polymers: microbe-host interactions illuminate the cytoskeleton
J Cell Biol.
.
Abstract
Intracellular pathogens subvert the host cell cytoskeleton to promote their own survival, replication, and dissemination. Study of these microbes has led to many discoveries about host cell biology, including the identification of cytoskeletal proteins, regulatory pathways, and mechanisms of cytoskeletal function. Actin is a common target of bacterial pathogens, but recent work also highlights the use of microtubules, cytoskeletal motors, intermediate filaments, and septins. The study of pathogen interactions with the cytoskeleton has illuminated key cellular processes such as phagocytosis, macropinocytosis, membrane trafficking, motility, autophagy, and signal transduction.
Figures
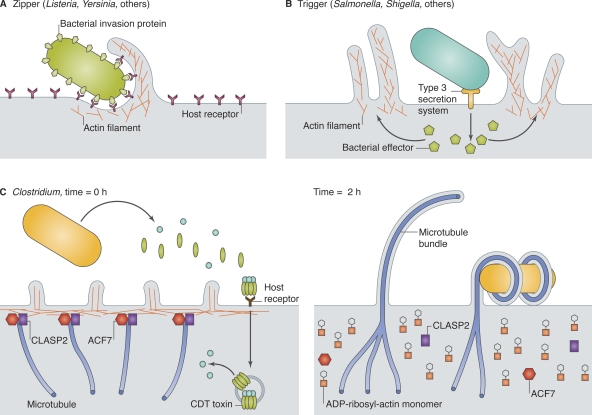
Bacteria exploit actin and microtubules to promote invasion and adherence. (A) Zippering bacteria express an invasion protein on their surface, which binds to a host receptor and initiates actin-dependent phagocytosis. (B) Triggering bacteria inject protein effector(s) across the host cell membrane, usually via a T3SS, leading to actin-dependent macropinocytosis. (C) Clostridium difficile transferase (CDT), a binary toxin, is endocytosed by intestinal epithelial cells, and the A subunit is released into the cytosol (left). CDT toxin ADP-ribosylates actin, promoting actin filament disassembly, effacement of microvilli, and release of cortical proteins that normally capture and stabilize microtubules. Unrestrained microtubule growth produces cellular extensions that wrap around external bacteria.
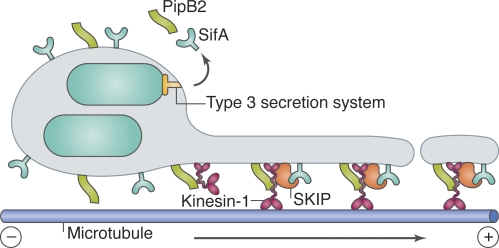
Model for kinesin-1 and SKIP activity on SCV membranes. The T3SS-secreted Salmonella effectors PipB2 and SifA recruit kinesin-1 and SKIP, respectively, to SCV membranes. It has been proposed that SKIP might then activate kinesin-1 by binding to the kinesin light chain, and that SKIP–kinesin-1 complexes promote tubulation and/or scission of SCV-derived membrane compartments, which are transported toward the plus end of microtubules.
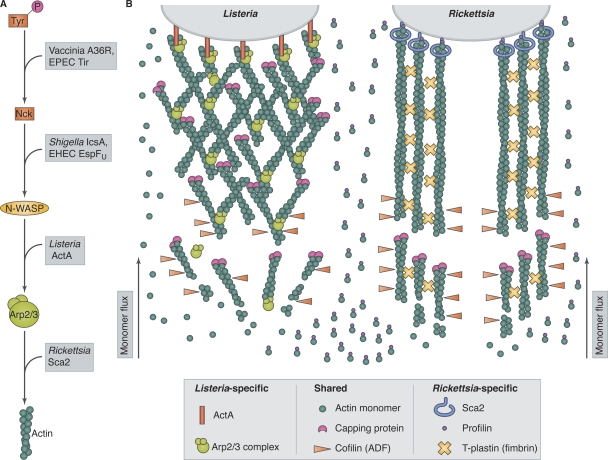
Pathogens use distinct actin-based motility mechanisms. (A) Pathogens intercept actin assembly pathways at different levels. Vaccinia virus and EPEC express proteins (A36R and Tir) that mimic host phosphotyrosine motifs to recruit the adaptor protein Nck. Shigella and EHEC produce proteins (IcsA and EspFU) that recruit and activate N-WASP. Listeria ActA mimics N-WASP to activate the host Arp2/3 complex. Rickettsia bypasses host nucleators using the formin-like protein Sca2 to interact directly with actin. (B) The actin-based motilities of Listeria and Rickettsia have distinct host protein requirements. Listeria expresses ActA on its surface, which activates the host Arp2/3 complex, producing branched actin filaments. Actin monomers or profilin–actin complexes can polymerize onto filament ends in Listeria tails. Rickettsia expresses the formin-like protein Sca2 on its surface, which nucleates unbranched actin filaments and requires profilin for filament elongation. T-plastin is also important for Rickettsia tail formation. Both systems require capping protein and cofilin (ADF).
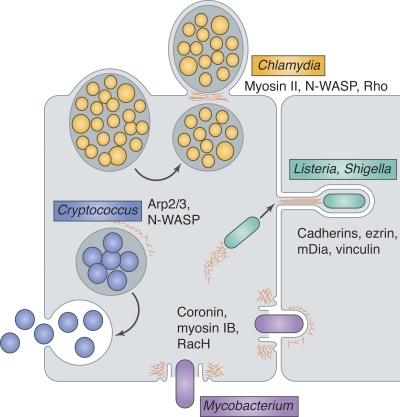
Exit strategies of diverse intracellular pathogens use actin.
Chlamydia (yellow) vacuoles are extruded through a cortical constriction, and the plasma membrane seals around the constriction point in a manner dependent on actin (red), releasing a double-membrane–bound bacterial compartment. The extrusion pathway appears to require myosin II and N-WASP for initiation and Rho for detachment of the extruded vacuole from the host cell. Listeria and Shigella (green), propelled by actin-based motility, enter plasma membrane protrusions and are taken up by neighboring cells. Cadherins, ezrin, mDia, and vinculin have been implicated in protrusion formation. Mycobacterium (purple) exits cells through a plasma membrane break surrounded by a barrel-shaped ejectosome rich in actin, myosin IB, and coronin. In host cells lacking RacH, ejectosomes are not detected and mycobacterial spreading is impaired. Cryptococcus (blue) phagosomes fuse with the plasma membrane, and intermittent actin polymerization around the phagosome, apparently mediated by N-WASP and the Arp2/3 complex, inhibits this fusion.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
