

Neural circuitry and plasticity mechanisms underlying delay eyeblink conditioning
- PMID: 21969489
- PMCID: PMC3861981
- DOI: 10.1101/lm.2023011
Neural circuitry and plasticity mechanisms underlying delay eyeblink conditioning
Abstract
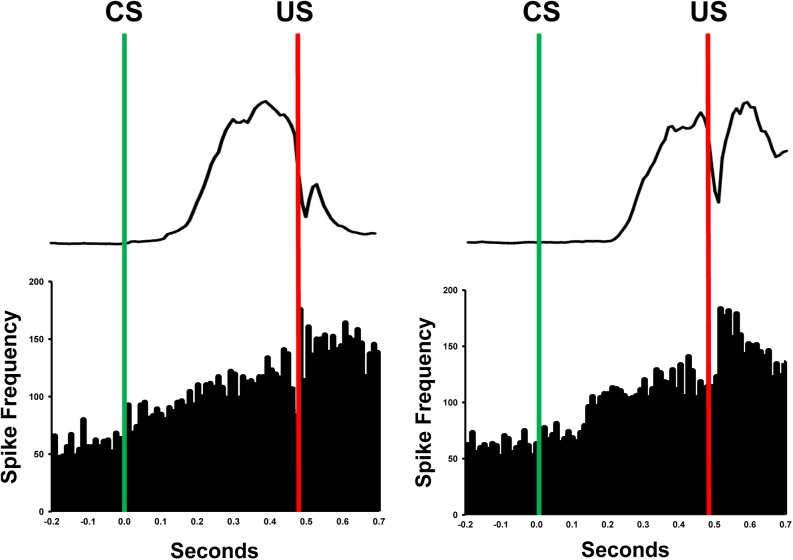
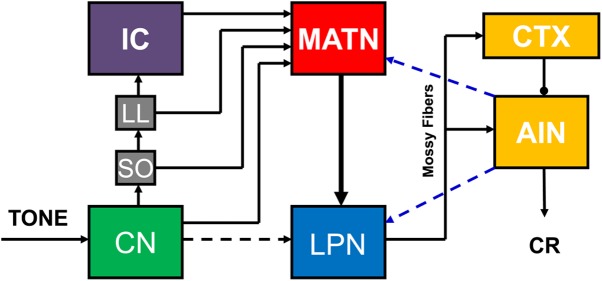
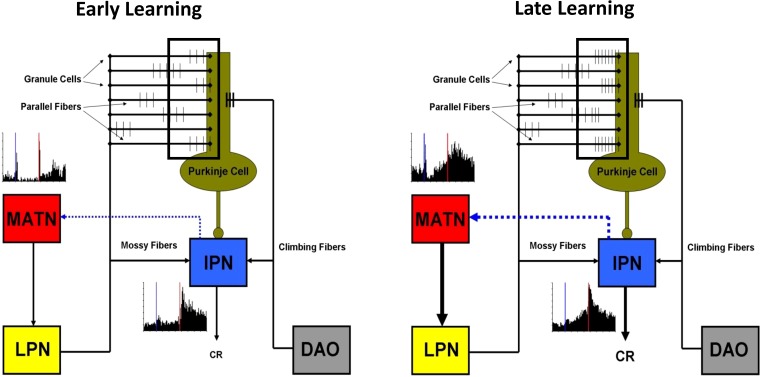
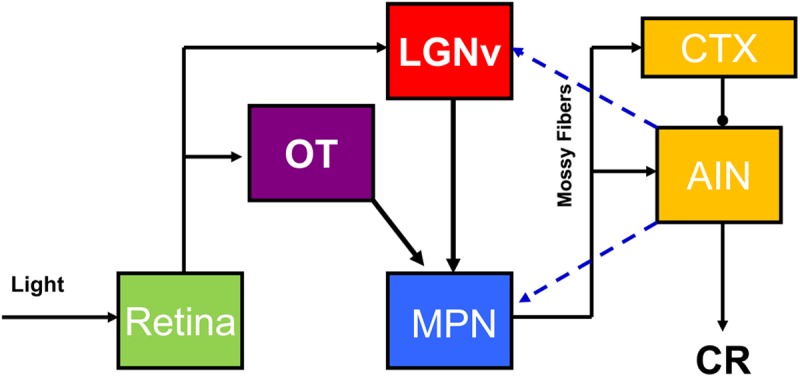
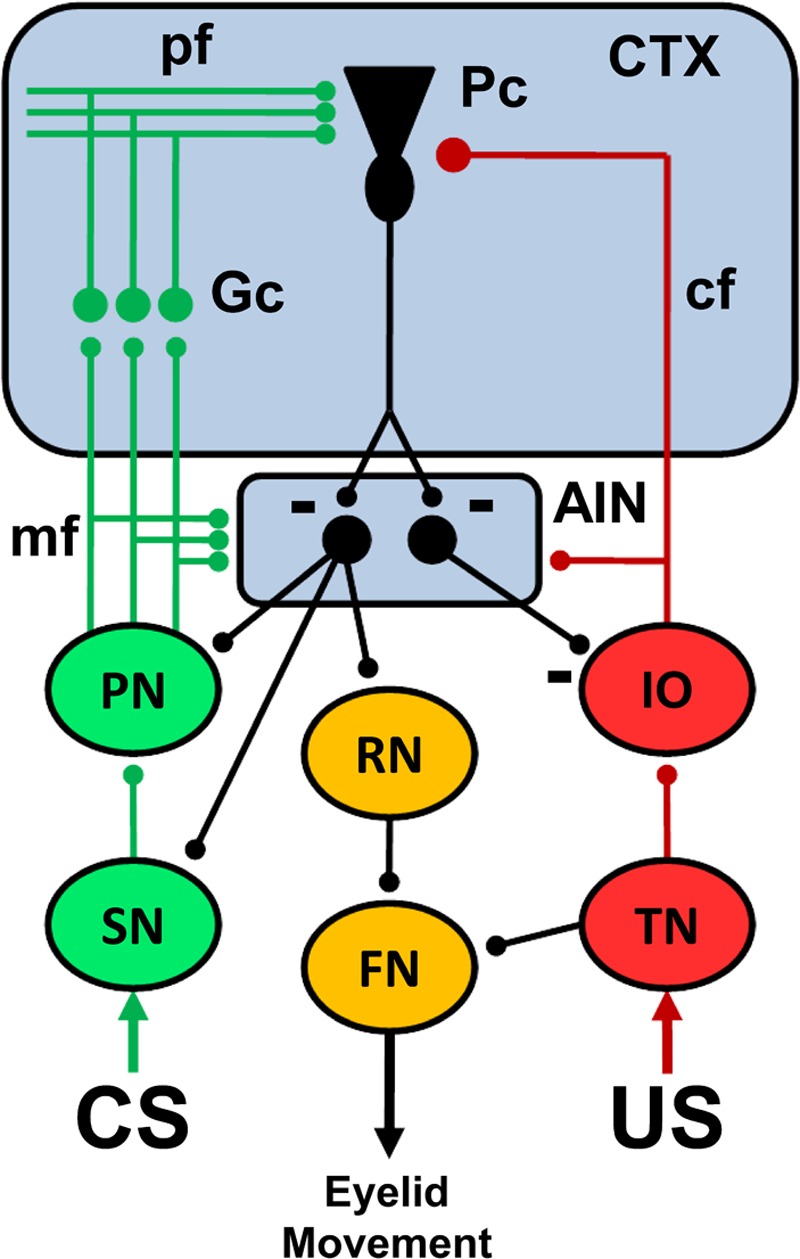
Pavlovian eyeblink conditioning has been used extensively as a model system for examining the neural mechanisms underlying associative learning. Delay eyeblink conditioning depends on the intermediate cerebellum ipsilateral to the conditioned eye. Evidence favors a two-site plasticity model within the cerebellum with long-term depression of parallel fiber synapses on Purkinje cells and long-term potentiation of mossy fiber synapses on neurons in the anterior interpositus nucleus. Conditioned stimulus and unconditioned stimulus inputs arise from the pontine nuclei and inferior olive, respectively, converging in the cerebellar cortex and deep nuclei. Projections from subcortical sensory nuclei to the pontine nuclei that are necessary for eyeblink conditioning are beginning to be identified, and recent studies indicate that there are dynamic interactions between sensory thalamic nuclei and the cerebellum during eyeblink conditioning. Cerebellar output is projected to the magnocellular red nucleus and then to the motor nuclei that generate the blink response(s). Tremendous progress has been made toward determining the neural mechanisms of delay eyeblink conditioning but there are still significant gaps in our understanding of the necessary neural circuitry and plasticity mechanisms underlying cerebellar learning.
Figures
References
-
- Aiba A, Kano M, Chen C, Stanton ME, Fox GD, Herrup K, Zwingman TA, Tonegawa S 1994. Deficient cerebellar long-term depression and impaired motor learning in mGluR1 mutant mice. Cell 79: 377–388 - PubMed
-
- Aksenov D, Serdyukova N, Irwin K, Bracha V 2004. GABA neurotransmission in the cerebellar interposed nuclei: Involvement in classically conditioned eyeblinks and neuronal activity. J Neurophysiol 91: 719–727 - PubMed
-
- Albus JS 1971. A theory of cerebellar function. Math Biosci 10: 25–61
-
- Allen MT, Padilla Y, Gluck MA 2002. Ibotenic acid lesions of the medial septum retard delay eyeblink conditioning in rabbits (Oryctolagus cuniculus). Behav Neurosci 116: 733–738 - PubMed
-
- Andersson G, Garwicz M, Hesslow G 1988. Evidence for a GABA-mediated cerebellar inhibition of the inferior olive in the cat. Exp Brain Res 72: 450–456 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources