

Allosteric vs. spontaneous exit-site (E-site) tRNA dissociation early in protein synthesis
- PMID: 21969541
- PMCID: PMC3193197
- DOI: 10.1073/pnas.1106999108
Allosteric vs. spontaneous exit-site (E-site) tRNA dissociation early in protein synthesis
Abstract
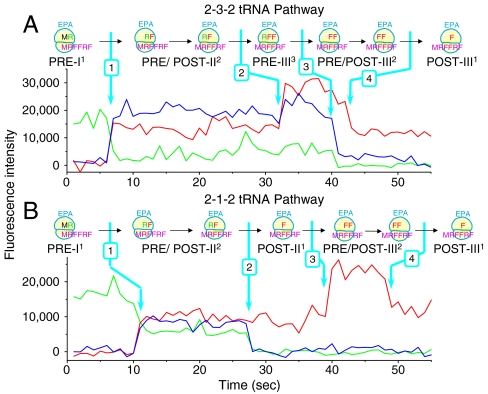
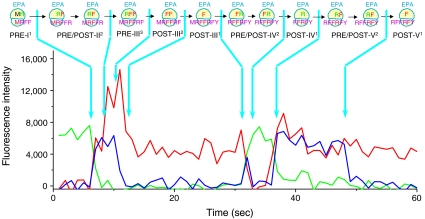
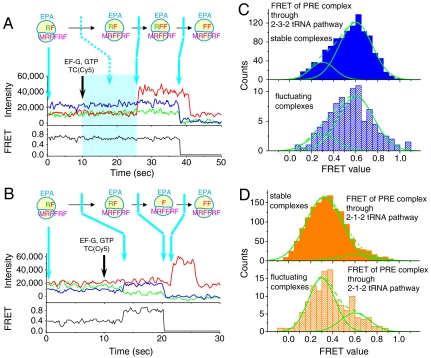
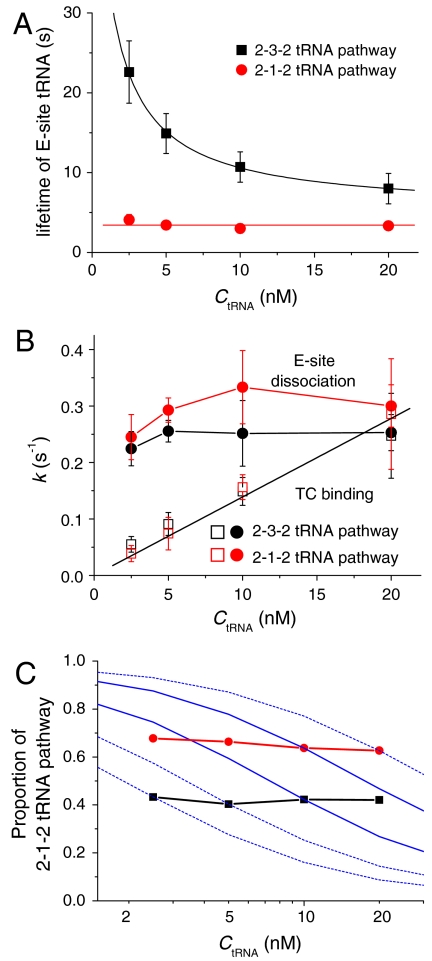
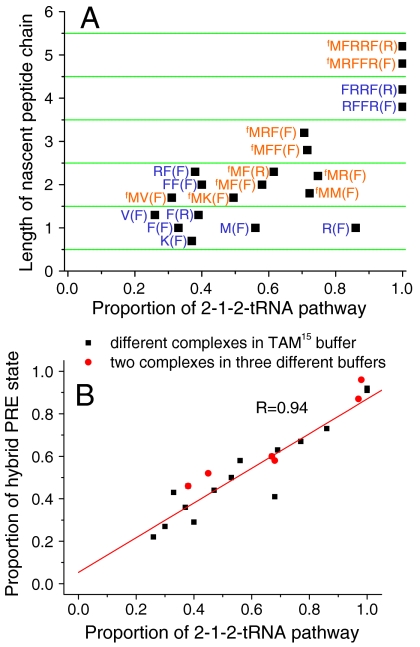
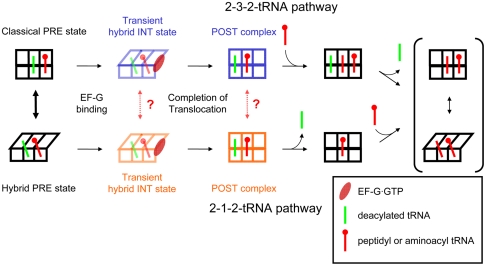
During protein synthesis, deacylated transfer RNAs leave the ribosome via an exit (E) site after mRNA translocation. How the ribosome regulates tRNA dissociation and whether functional linkages between the aminoacyl (A) and E sites modulate the dynamics of protein synthesis have long been debated. Using single molecule fluorescence resonance energy transfer experiments, we find that, during early cycles of protein elongation, tRNAs are often held in the E site until being allosterically released when the next aminoacyl tRNA binds to the A site. This process is regulated by the length and sequence of the nascent peptide and by the conformational state, detected by tRNA proximity, prior to translocation. In later cycles, E-site tRNA dissociates spontaneously. Our results suggest that the distribution of pretranslocation tRNA states and posttranslocation pathways are correlated within each elongation cycle via communication between distant subdomains in the ribosome, but that this correlation between elongation cycle intermediates does not persist into succeeding cycles.
Conflict of interest statement
Conflict of interest statement: B.S.C. and Y.E.G. are paid consultants of Anima Cell Metrology, Inc.
Figures
References
-
- Schmeing TM, Ramakrishnan V. What recent ribosome structures have revealed about the mechanism of translation. Nature. 2009;461:1234–1242. - PubMed
-
- Ramakrishnan V. Ribosome structure and the mechanism of translation. Cell. 2002;108:557–572. - PubMed
-
- Mitra K, Frank J. Ribosome dynamics: Insights from atomic structure modeling into cryo-electron microscopy maps. Annu Rev Biophys Biomol Struct. 2006;35:299–317. - PubMed
-
- Moazed D, Noller HF. Intermediate states in the movement of transfer-RNA in the ribosome. Nature. 1989;342:142–148. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
