

Positive feedback between PGE2 and COX2 redirects the differentiation of human dendritic cells toward stable myeloid-derived suppressor cells
- PMID: 21972293
- PMCID: PMC3217352
- DOI: 10.1182/blood-2011-07-365825
Positive feedback between PGE2 and COX2 redirects the differentiation of human dendritic cells toward stable myeloid-derived suppressor cells
Abstract
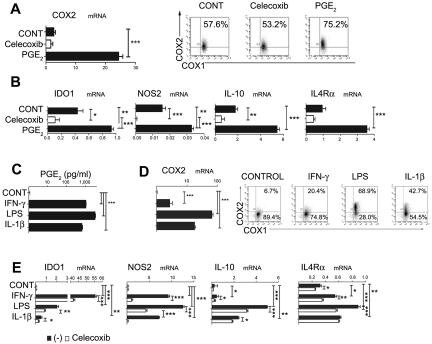
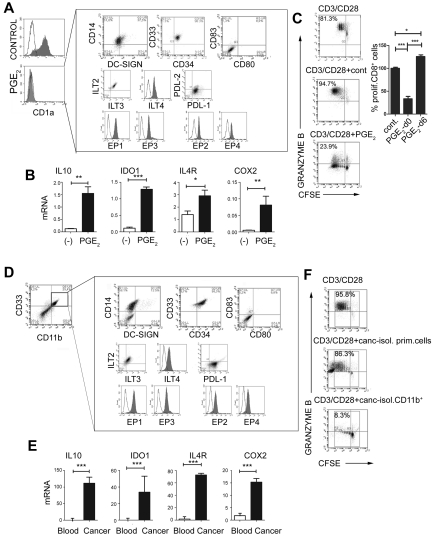
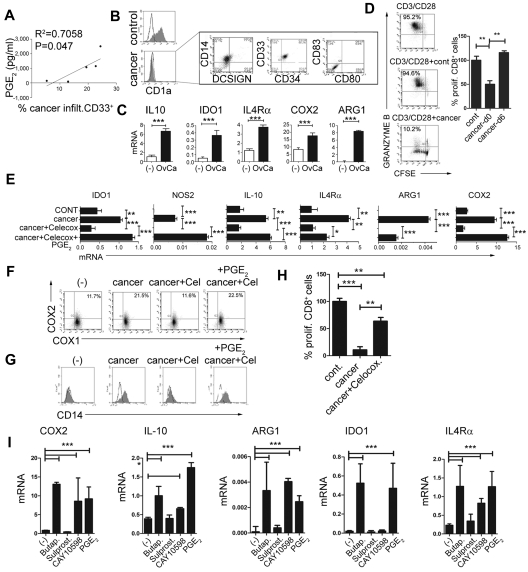
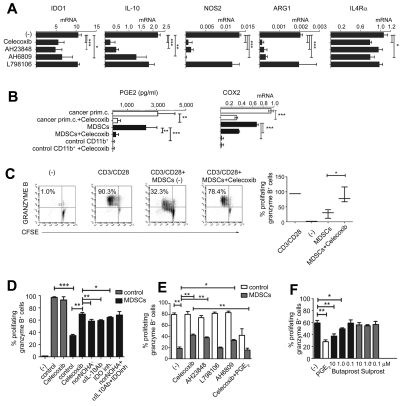
Dendritic cells (DCs) and myeloid-derived suppressor cells (MDSCs) show opposing roles in the immune system. In the present study, we report that the establishment of a positive feedback loop between prostaglandin E(2) (PGE(2)) and cyclooxygenase 2 (COX2), the key regulator of PGE(2) synthesis, represents the determining factor in redirecting the development of CD1a(+) DCs to CD14(+)CD33(+)CD34(+) monocytic MDSCs. Exogenous PGE(2) and such diverse COX2 activators as lipopolysaccharide, IL-1β, and IFNγ all induce monocyte expression of COX2, blocking their differentiation into CD1a(+) DCs and inducing endogenous PGE(2), IDO1, IL-4Rα, NOS2, and IL-10, typical MDSC-associated suppressive factors. The addition of PGE(2) to GM-CSF/IL-4-supplemented monocyte cultures is sufficient to induce the MDSC phenotype and cytotoxic T lymphocyte (CTL)-suppressive function. In accordance with the key role of PGE(2) in the physiologic induction of human MDSCs, the frequencies of CD11b(+)CD33(+) MDSCs in ovarian cancer are closely correlated with local PGE(2) production, whereas the cancer-promoted induction of MDSCs is strictly COX2 dependent. The disruption of COX2-PGE(2) feedback using COX2 inhibitors or EP2 and EP4 antagonists suppresses the production of MDSC-associated suppressive factors and the CTL-inhibitory function of fully developed MDSCs from cancer patients. The central role of COX2-PGE(2) feedback in the induction and persistence of MDSCs highlights the potential for its manipulation to enhance or suppress immune responses in cancer, autoimmunity, or transplantation.
Figures
References
-
- Pulendran B, Palucka K, Banchereau J. Sensing pathogens and tuning immune responses. Science. 2001;293(5528):253–256. - PubMed
-
- Torres-Aguilar H, Blank M, Jara LJ, Shoenfeld Y. Tolerogenic dendritic cells in autoimmune diseases: crucial players in induction and prevention of autoimmunity. Autoimmun Rev. 2010;10(1):8–17. - PubMed
-
- Pulendran B, Tang H, Manicassamy S. Programming dendritic cells to induce T(H)2 and tolerogenic responses. Nat Immunol. 2010;11(8):647–655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
