

doi: 10.1101/gad.17224711.
TNF-stimulated MAP kinase activation mediated by a Rho family GTPase signaling pathway
Affiliations
- PMID: 21979919
- PMCID: PMC3197205
- DOI: 10.1101/gad.17224711
Item in Clipboard
TNF-stimulated MAP kinase activation mediated by a Rho family GTPase signaling pathway
Genes Dev.
.
Abstract
The biological response to tumor necrosis factor (TNF) involves activation of MAP kinases. Here we report a mechanism of MAP kinase activation by TNF that is mediated by the Rho GTPase family members Rac/Cdc42. This signaling pathway requires Src-dependent activation of the guanosine nucleotide exchange factor Vav, activation of Rac/Cdc42, and the engagement of the Rac/Cdc42 interaction site (CRIB motif) on mixed-lineage protein kinases (MLKs). We show that this pathway is essential for full MAP kinase activation during the response to TNF. Moreover, this MLK pathway contributes to inflammation in vivo.
Figures
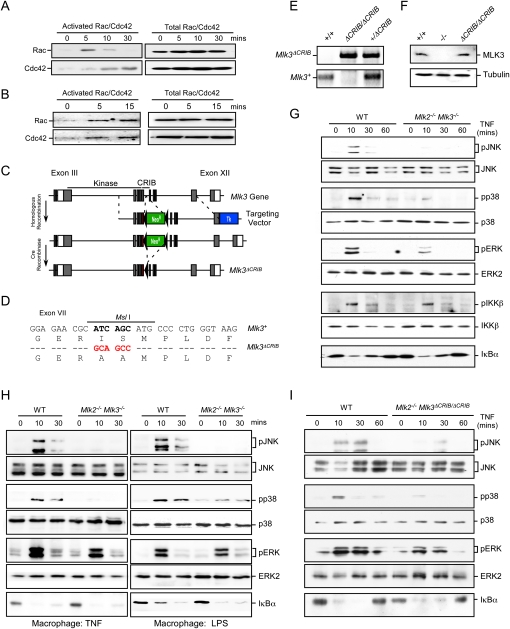
TNF causes activation of a Rac/Cdc42–MLK signal transduction pathway. (A,B) MEFs (A) or BMDMs (B) were treated without or with 10 ng/mL TNFα. Activated (GTP-loaded) Rac/Cdc42 was isolated and quantitated by immunoblot analysis. The amount of Rac/Cdc42 in cell lysates was measured by immunoblot analysis. (C) The structure of the Mlk3 genomic locus and the targeting vector are illustrated. Homologous recombination causes the replacement of Mlk3 exon VII with a mutated form of exon VII together with the insertion of a floxed NeoR cassette. The floxed NeoR cassette was excised using Cre recombinase. (D) The point mutations in exon VII created by the targeting vector are illustrated. The nucleotide changes cause the elimination of an MslI restriction site and the introduction of two amino acid substitutions (Ile492Ala and Ser493Ala) that disrupt the MLK3 CRIB domain. (E) Genomic DNA from wild-type, Mlk3+/ΔCRIB, and Mlk3ΔCRIB/ΔCRIB mice was examined by PCR analysis to detect the wild-type and mutant Mlk3 alleles. (F) Extracts prepared from wild-type (WT), Mlk3−/−, and Mlk3ΔCRIB/ΔCRIB MEFs were examined by immunoblot analysis with antibodies to MLK3 and α-Tubulin. (G–I) Wild-type and Mlk2−/− Mlk3−/− MEFs (G), wild-type and Mlk2−/− Mlk3−/− BMDMs (H), or wild-type and Mlk2−/− Mlk3ΔCRIB/ ΔCRIB MEFs (I) were treated without or with 10 ng/mL TNF or 100 ng/mL LPS. MAP kinase and IKKβ activation and the degradation of IκBα were examined by immunoblot analysis.
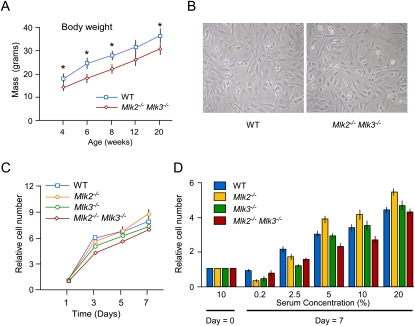
The effect of MLK2 and MLK3 deficiency on growth. (A) The body mass of male wild-type (WT) and Mlk2−/− Mlk3−/− mice was measured at different time intervals (mean ± SD; n = 8). Statistically significant differences are indicated. (*) P < 0.05. (B) Wild-type and Mlk2−/− Mlk3−/− MEFs were examined by phase-contrast microscopy. (C) The proliferation of wild-type, Mlk2−/−, Mlk3−/−, and Mlk2−/− Mlk3−/− MEFs during culture for 7 d was examined by plating 2.5 × 104 MEFs in 11-mm wells using Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum. Relative cell numbers were measured by staining with crystal violet (mean ± SD; n = 6). No significant differences between wild-type and MLK-deficient MEFs were detected. P > 0.05. (D) MEFs were incubated for 0 or 7 d in Dulbecco's modified Eagle's medium supplemented with different amounts of fetal bovine serum. Relative cell numbers were measured by staining with crystal violet (mean ± SD; n = 6). No significant differences between wild-type and MLK-deficient MEFs were detected. P > 0.05.
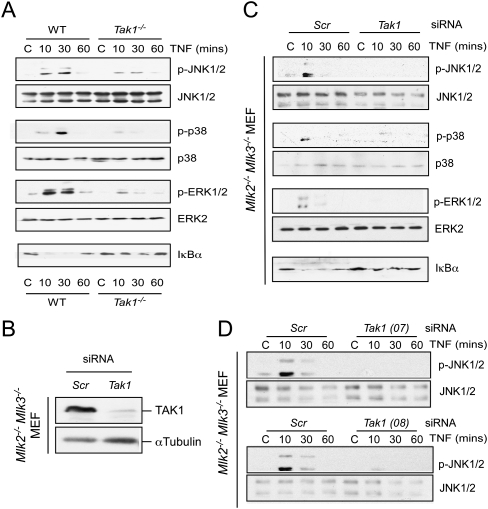
Residual MAP kinase activation in MLK-deficient MEFs is mediated by TAK1. (A) Wild-type (WT) and Tak1−/− MEFs were treated without or with TNFα (10 ng/mL). The activation of MAP kinases and the degradation of IκBα was examined by immunoblot analysis. (B,C) Mlk2−/− Mlk3−/− MEFs were transfected with control (Scr) siRNA or Tak1 siRNA (72 h). Transfection assays were performed using DharmaFECT 3 reagent with ON-TARGET plus Nontargeting Pool D-001810-10 for control or ON-TARGET plus SMART pool L-040718-00 for Tak1 (MAP3K7) siRNA (Thermo Scientific). (B) The expression of TAK1 and α-Tubulin was examined by immunoblot analysis. (C) MAP kinase activation and the degradation of IκBα in response to treatment with TNFα (10 ng/mL) were examined by immunoblot analysis. (D) Mlk2−/− Mlk3−/− MEFs were transfected with scrambled (Scr) siRNA or Tak1 siRNA (72 h). The MEFs were treated without or with 10 ng/mL TNFα. Protein extracts were examined by immunoblot analysis by probing with antibodies to JNK1/2 and phospho-JNK. The siRNA for Tak1 were ON-TARGET plus siRNA J-040718-07 (top panel) and J-040718-08 (bottom panel).
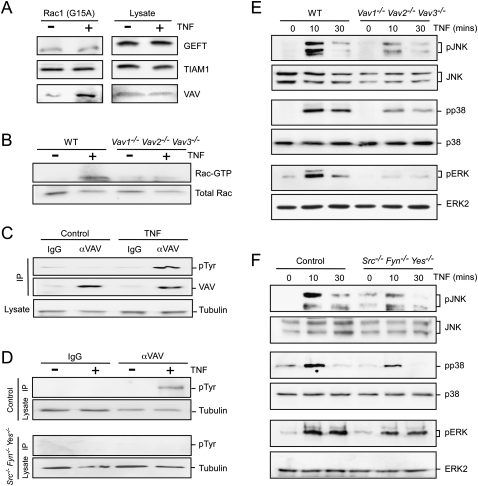
TNF-stimulated Rac activation is mediated by Src and Vav. (A) Lysates prepared from BMDMs treated without or with 10 ng/mL TNF (15 min) were incubated with immobilized Rac1G15A. Bound proteins and the cell lysates were examined by immunoblot analysis using antibodies to GEFT, TIAM1, and Vav. (B) Wild-type (WT) and Vav1−/− Vav2−/− Vav3−/− BMDMs were treated without or with 10 ng/mL TNFα. Activated (GTP-loaded) Rac was isolated and quantitated by immunoblot analysis. The amount of Rac in cell lysates was measured by immunoblot analysis. (C) Lysates prepared from wild-type BMDMs treated without or with TNF were examined by immunoprecipitation with a control antibody (IgG) or with an antibody to Vav. The immunoprecipitates were examined by immunoblot analysis using antibodies to phosphotyrosine (pTyr) and Vav. The cell lysates were examined by probing with an antibody to α-Tubulin. (D) Src−/− Fyn−/− Yes−/− fibroblasts complemented with Src (Control) or without Src were treated without or with TNFα. Lysates were examined by immunoprecipitation with a control antibody (IgG) or with an antibody to Vav. The immunoprecipitates were examined by immunoblot analysis using antibodies to pTyr. The cell lysates were examined by probing with an antibody to α-Tubulin. (E,F) Wild-type and Vav1−/− Vav2−/− Vav3−/− BMDMs (E) or Control and Src−/− Fyn−/− Yes−/− fibroblasts (F) were treated without or with 10 ng/mL TNF. MAP kinase activation was examined by immunoblot analysis.
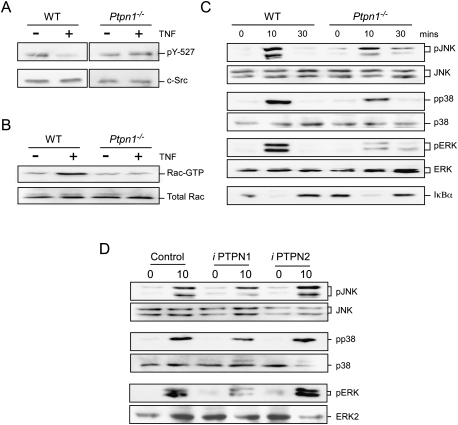
The protein tyrosine phosphatase PTPN1 contributes to TNF-stimulated MAP kinase activation. (A) Lysates prepared from wild-type (WT) and Ptpn1−/− BMDMs were treated without or with 10 ng/mL TNFα and examined by immunoblot analysis using antibodies to Src and pY527 Src. (B) Wild-type and Ptpn1−/− BMDMs were treated without or with 10 ng/mL TNF (15 min). Activated (GTP-loaded) Rac1 was isolated and quantitated by immunoblot analysis. The amount of Rac1 in cell lysates was measured by immunoblot analysis. (C) Wild-type and Ptpn1−/− BMDMs were treated without or with 10 ng/mL TNF. MAP kinase activation and the degradation of IκBα was examined by immunoblot analysis. (D) BMDMs were treated with solvent or with drugs that selectively inhibit PTPN1 and PTPN2 (30 min). The cells were then treated with or without 10 ng/mL TNFα (10 min). Cell lysates were examined by immunoblot analysis using antibodies to phospho-JNK, JNK, phospho-p38, p38α, phospho-ERK, and ERK2.
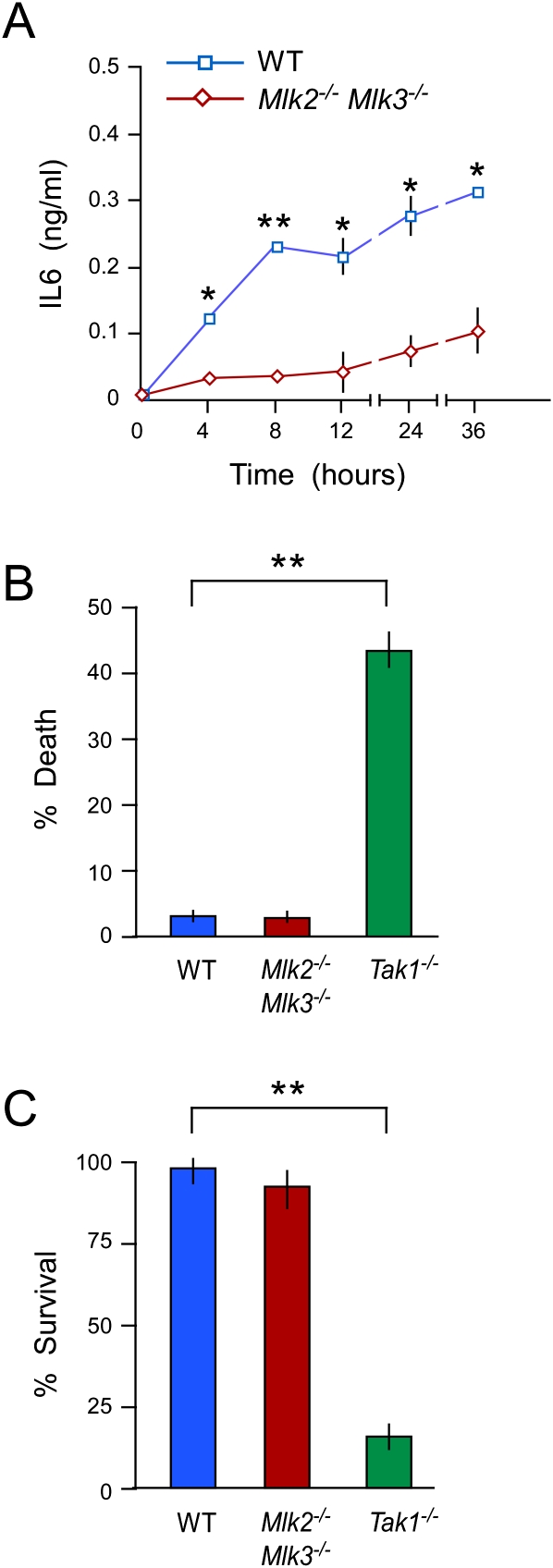
The MLK pathway contributes to TNF-stimulated signal transduction. (A) Wild-type (WT) and Mlk2−/− Mlk3−/− MEFs were treated with 10 ng/mL TNFα. The amount of IL6 in the culture medium was measured by ELISA (mean ± SD; n = 6). Statistically significant differences between wild-type and Mlk2−/− Mlk3−/− MEFs are indicated. (*) P < 0.05; (**) P < 0.01. (B,C) Wild-type, Mlk2−/− Mlk3−/−, and Tak1−/− MEFs were treated with 10 ng/mL TNF (24 h). Cell death was examined by measuring the presence of lactate dehydrogenase in the medium (B), and cell viability was measured by staining with crystal violet (C). The data presented are the mean ± SD (n = 4). Statistically significant differences between wild-type and Tak1−/− MEFs are indicated. (**) P < 0.01. No statistically significant differences between wild-type and Mlk2−/− Mlk3−/− MEFs were detected.
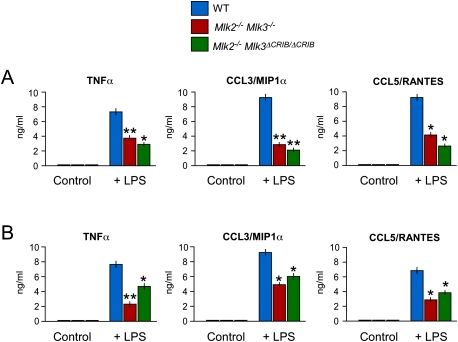
The MLK pathway contributes to LPS-mediated inflammation. (A) Wild-type (WT) mice, Mlk2−/− Mlk3−/− mice, and Mlk2−/− Mlk3ΔCRIB/ΔCRIB mice were treated without and with 20 mg/kg LPS by intraperitoneal injection (2 h). The amount of TNFα, CCL3/MIPα, and CCL5/RANTES in the serum was measured by ELISA (mean ± SD; n = 8). Statistically significant differences between wild-type and MLK-deficient mice are indicated. (*) P < 0.05; (**) P < 0.01. (B) Primary BMDMs were isolated from wild-type mice, Mlk2−/− Mlk3−/− mice, and Mlk2−/− Mlk3ΔCRIB/ΔCRIB mice. The BMDMs were treated without and with 100 ng/mL LPS (24 h). The amount of TNFα, CCL3/MIPα, and CCL5/RANTES in the culture medium was measured by ELISA (mean ± SD; n = 6). Statistically significant differences between wild-type and MLK-deficient BMDMs are indicated. (*) P < 0.05; (**) P < 0.01.
References
-
- Abe K, Rossman KL, Liu B, Ritola KD, Chiang D, Campbell SL, Burridge K, Der CJ 2000. Vav2 is an activator of Cdc42, Rac1, and RhoA. J Biol Chem 275: 10141–10149 - PubMed
-
- Bock BC, Vacratsis PO, Qamirani E, Gallo KA 2000. Cdc42-induced activation of the mixed-lineage kinase SPRK in vivo. Requirement of the Cdc42/Rac interactive binding motif and changes in phosphorylation. J Biol Chem 275: 14231–14241 - PubMed
-
- Chadee DN, Kyriakis JM 2004. MLK3 is required for mitogen activation of B-Raf, ERK and cell proliferation. Nat Cell Biol 6: 770–776 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous