

Monalysin, a novel ß-pore-forming toxin from the Drosophila pathogen Pseudomonas entomophila, contributes to host intestinal damage and lethality
- PMID: 21980286
- PMCID: PMC3182943
- DOI: 10.1371/journal.ppat.1002259
Monalysin, a novel ß-pore-forming toxin from the Drosophila pathogen Pseudomonas entomophila, contributes to host intestinal damage and lethality
Abstract
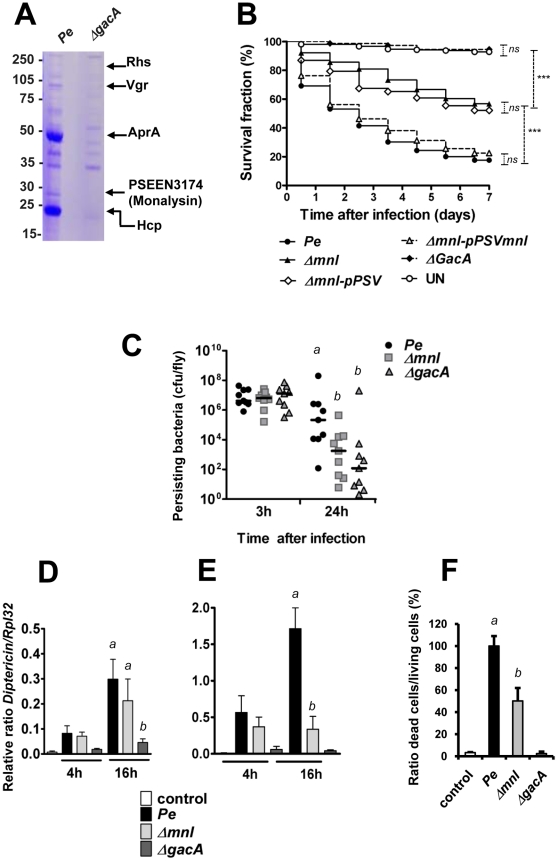
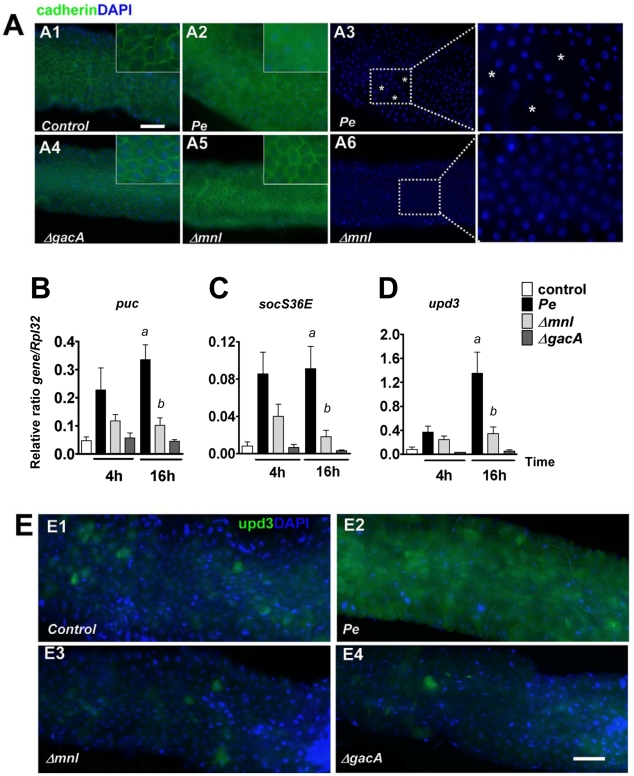
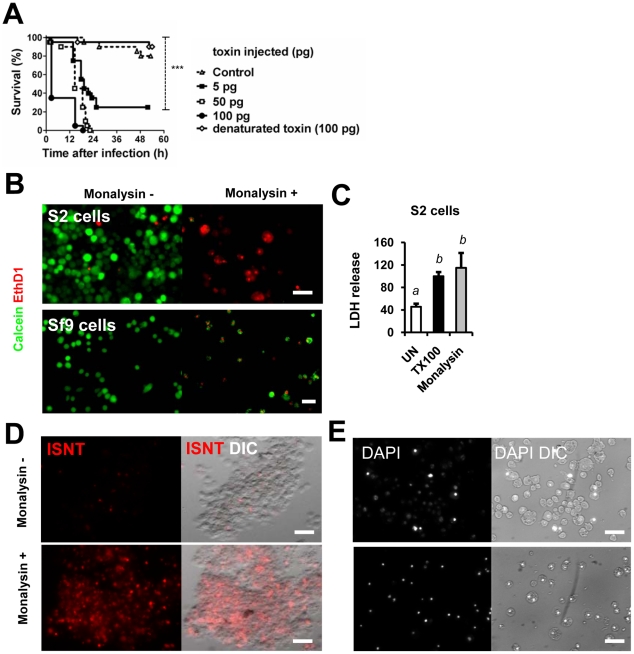
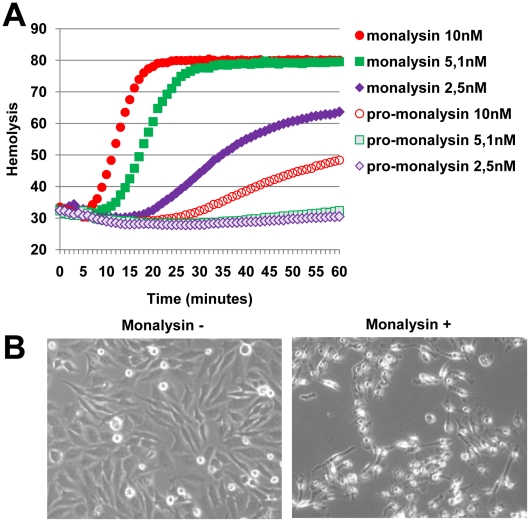
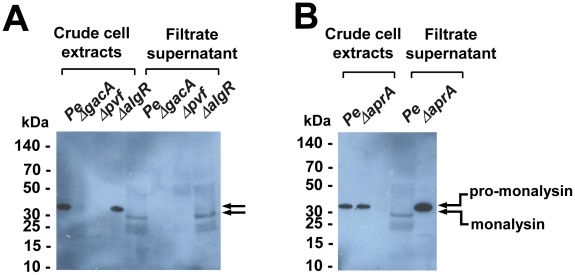
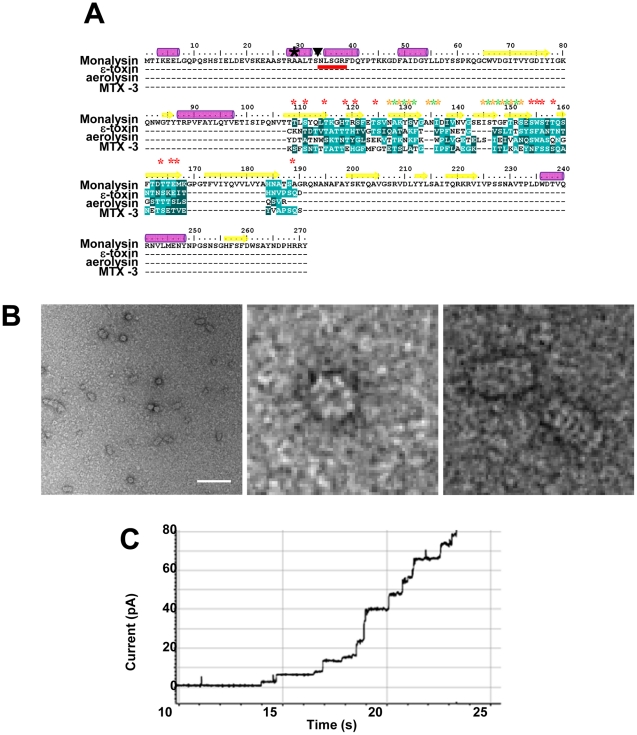
Pseudomonas entomophila is an entomopathogenic bacterium that infects and kills Drosophila. P. entomophila pathogenicity is linked to its ability to cause irreversible damages to the Drosophila gut, preventing epithelium renewal and repair. Here we report the identification of a novel pore-forming toxin (PFT), Monalysin, which contributes to the virulence of P. entomophila against Drosophila. Our data show that Monalysin requires N-terminal cleavage to become fully active, forms oligomers in vitro, and induces pore-formation in artificial lipid membranes. The prediction of the secondary structure of the membrane-spanning domain indicates that Monalysin is a PFT of the ß-type. The expression of Monalysin is regulated by both the GacS/GacA two-component system and the Pvf regulator, two signaling systems that control P. entomophila pathogenicity. In addition, AprA, a metallo-protease secreted by P. entomophila, can induce the rapid cleavage of pro-Monalysin into its active form. Reduced cell death is observed upon infection with a mutant deficient in Monalysin production showing that Monalysin plays a role in P. entomophila ability to induce intestinal cell damages, which is consistent with its activity as a PFT. Our study together with the well-established action of Bacillus thuringiensis Cry toxins suggests that production of PFTs is a common strategy of entomopathogens to disrupt insect gut homeostasis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Sansonetti PJ. War and peace at mucosal surfaces. Nat Rev Immunol. 2004;4:953–964. - PubMed
-
- Hooper LV, Macpherson AJ. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat Rev Immunol. 2010;10:159–169. - PubMed
-
- Vodovar N, Vallenet D, Cruveiller S, Rouy Z, Barbe V, et al. Complete genome sequence of the entomopathogenic and metabolically versatile soil bacterium Pseudomonas entomophila. Nat Biotechnol. 2006;24:673–679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
