

Inducing autophagy by rapamycin before, but not after, the formation of plaques and tangles ameliorates cognitive deficits
- PMID: 21980451
- PMCID: PMC3182203
- DOI: 10.1371/journal.pone.0025416
Inducing autophagy by rapamycin before, but not after, the formation of plaques and tangles ameliorates cognitive deficits
Abstract
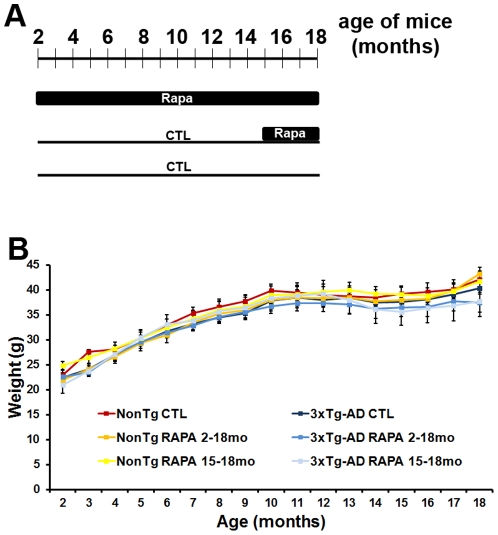
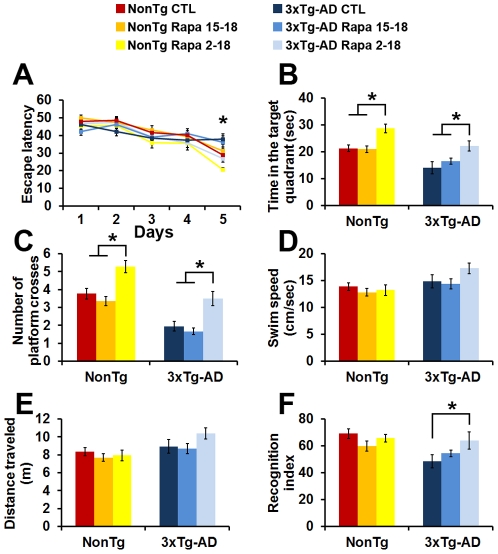
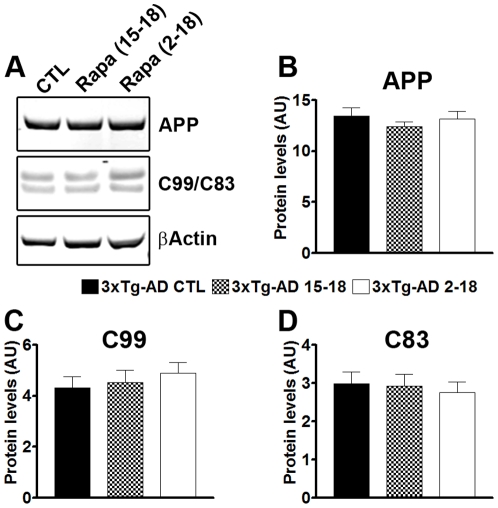
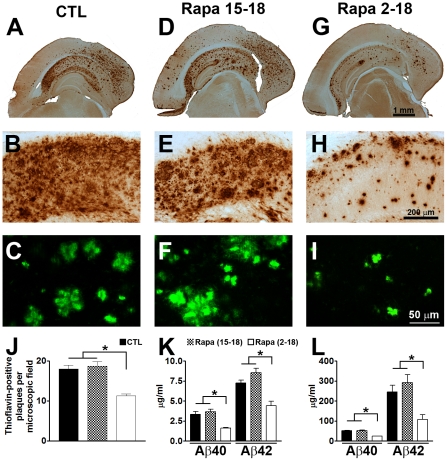
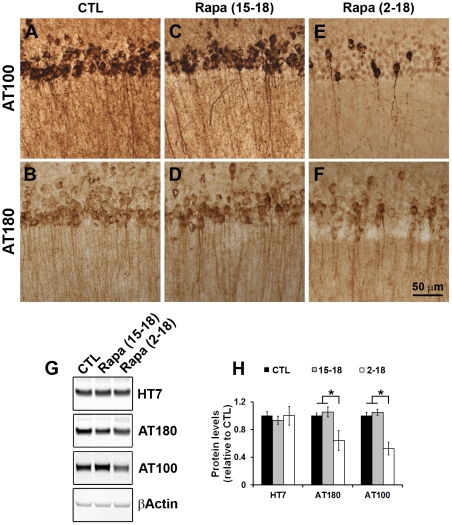
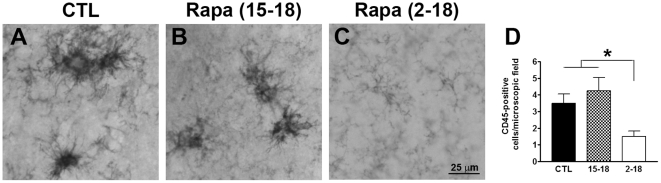
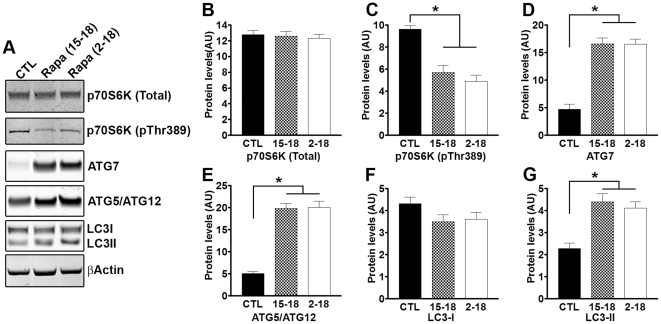
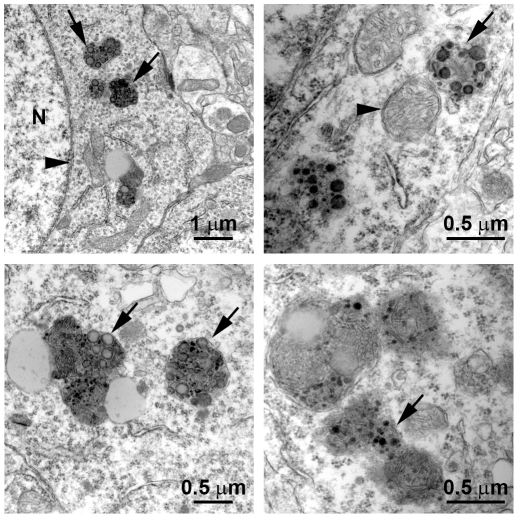
Previous studies have shown that inducing autophagy ameliorates early cognitive deficits associated with the build-up of soluble amyloid-β (Aβ). However, the effects of inducing autophagy on plaques and tangles are yet to be determined. While soluble Aβ and tau represent toxic species in Alzheimer's disease (AD) pathogenesis, there is well documented evidence that plaques and tangles also are detrimental to normal brain function. Thus, it is critical to assess the effects of inducing autophagy in an animal model with established plaques and tangles. Here we show that rapamycin, when given prophylactically to 2-month-old 3xTg-AD mice throughout their life, induces autophagy and significantly reduces plaques, tangles and cognitive deficits. In contrast, inducing autophagy in 15-month-old 3xTg-AD mice, which have established plaques and tangles, has no effects on AD-like pathology and cognitive deficits. In conclusion, we show that autophagy induction via rapamycin may represent a valid therapeutic strategy in AD when administered early in the disease progression.
Conflict of interest statement
Figures
References
-
- Querfurth HW, LaFerla FM. Alzheimer's disease. N Engl J Med. 2010;362:329–344. - PubMed
-
- Busche MA, Eichhoff G, Adelsberger H, Abramowski D, Wiederhold KH, et al. Clusters of hyperactive neurons near amyloid plaques in a mouse model of Alzheimer's disease. Science. 2008;321:1686–1689. - PubMed
-
- Ling D, Salvaterra PM. A central role for autophagy in Alzheimer-type neurodegeneration. Autophagy. 2009;5:738–740. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
