

Control of vocal and respiratory patterns in birdsong: dissection of forebrain and brainstem mechanisms using temperature
- PMID: 21980466
- PMCID: PMC3182229
- DOI: 10.1371/journal.pone.0025461
Control of vocal and respiratory patterns in birdsong: dissection of forebrain and brainstem mechanisms using temperature
Abstract
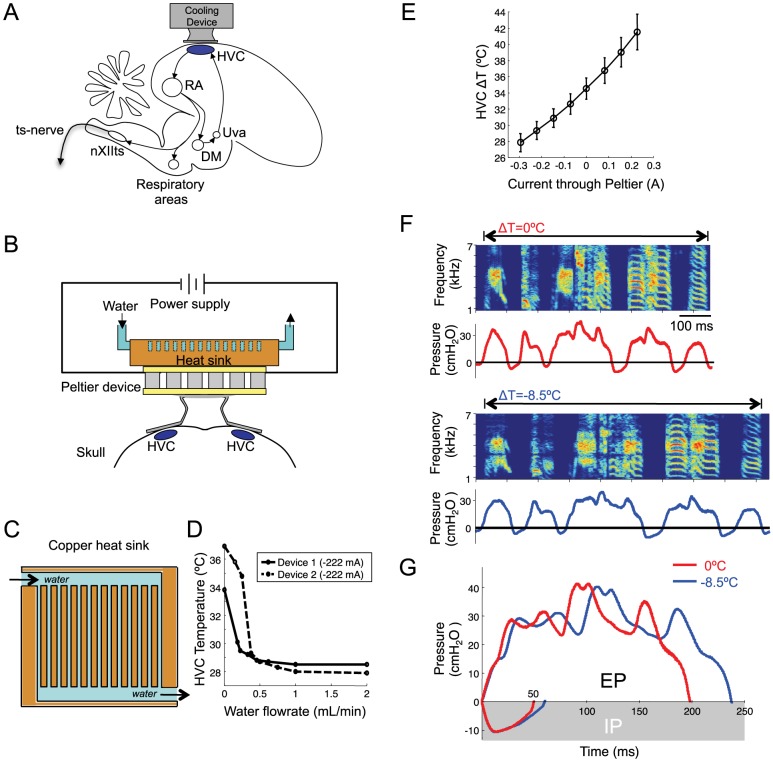
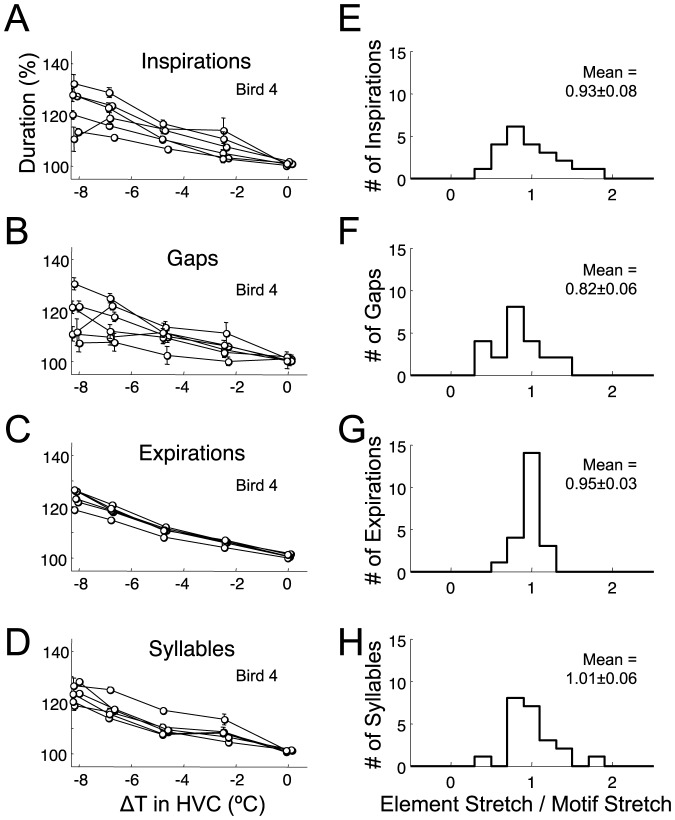
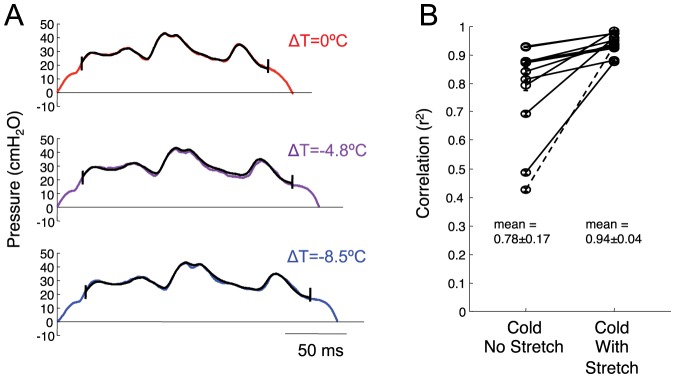
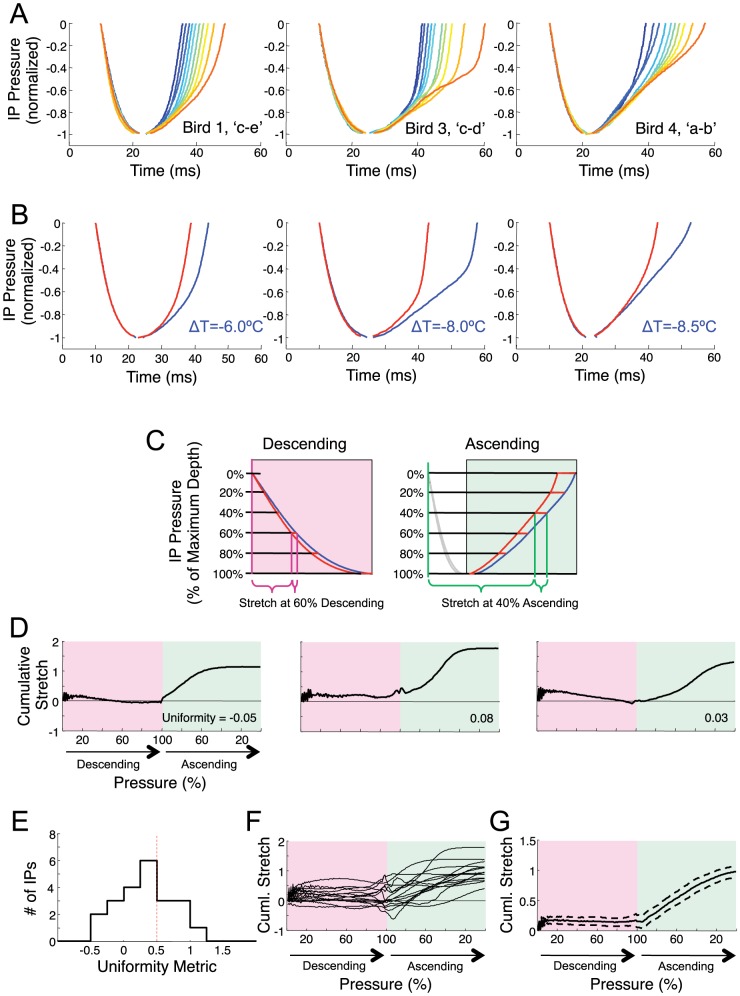
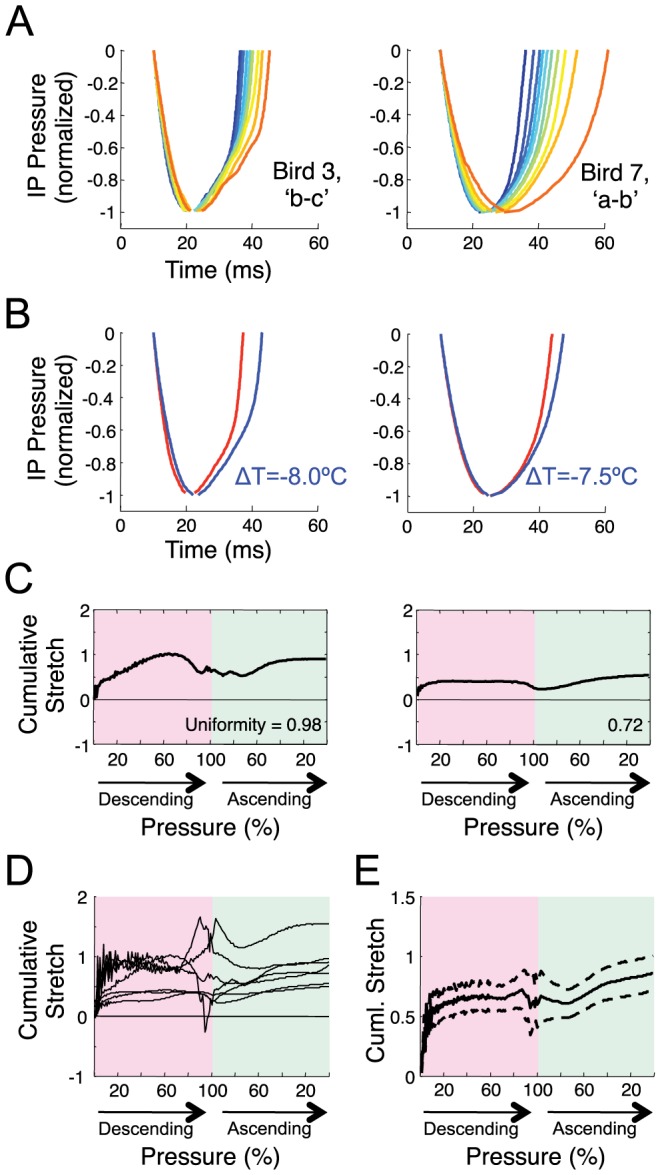
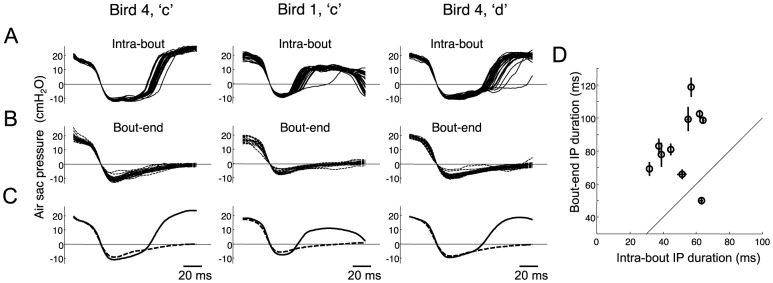
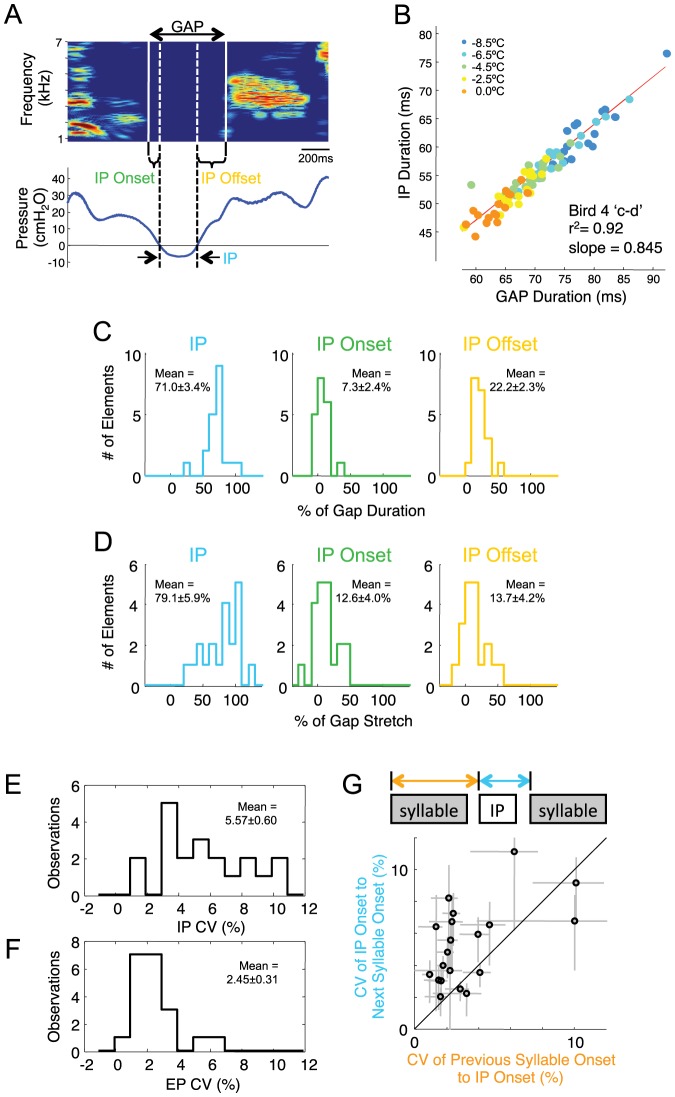
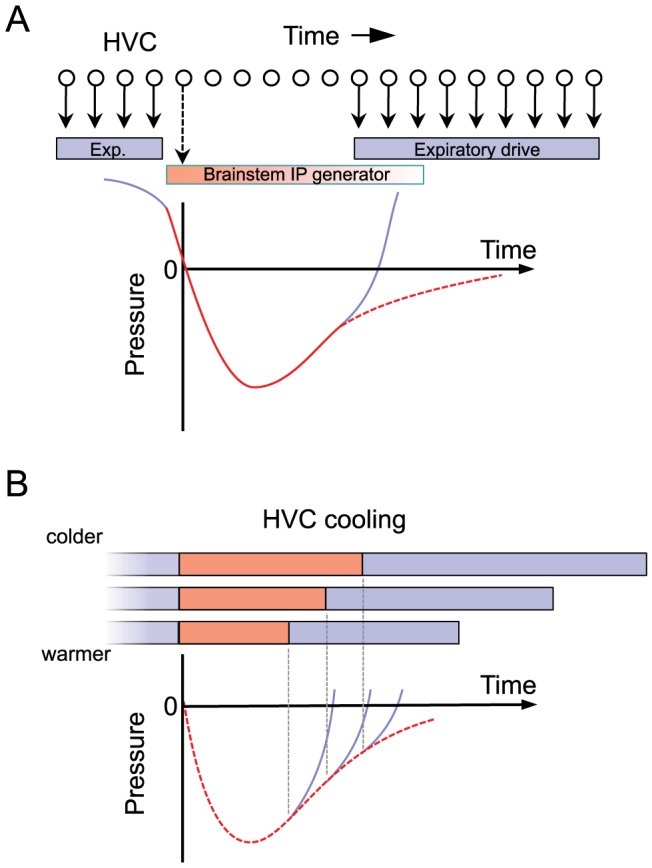
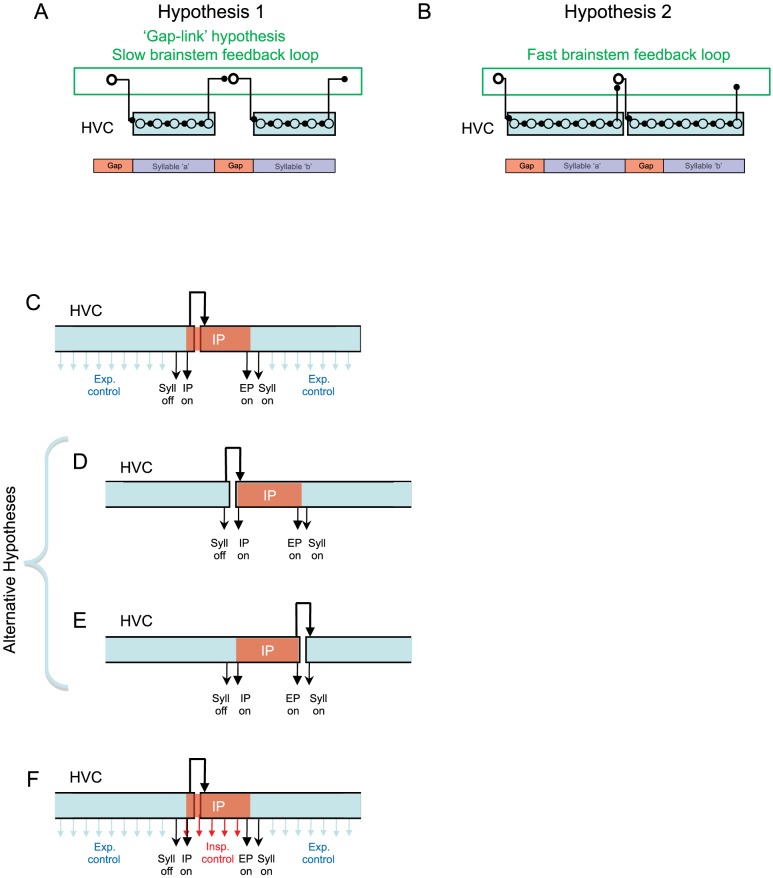
Learned motor behaviors require descending forebrain control to be coordinated with midbrain and brainstem motor systems. In songbirds, such as the zebra finch, regular breathing is controlled by brainstem centers, but when the adult songbird begins to sing, its breathing becomes tightly coordinated with forebrain-controlled vocalizations. The periods of silence (gaps) between song syllables are typically filled with brief breaths, allowing the bird to sing uninterrupted for many seconds. While substantial progress has been made in identifying the brain areas and pathways involved in vocal and respiratory control, it is not understood how respiratory and vocal control is coordinated by forebrain motor circuits. Here we combine a recently developed technique for localized brain cooling, together with recordings of thoracic air sac pressure, to examine the role of cortical premotor nucleus HVC (proper name) in respiratory-vocal coordination. We found that HVC cooling, in addition to slowing all song timescales as previously reported, also increased the duration of expiratory pulses (EPs) and inspiratory pulses (IPs). Expiratory pulses, like song syllables, were stretched uniformly by HVC cooling, but most inspiratory pulses exhibited non-uniform stretch of pressure waveform such that the majority of stretch occurred late in the IP. Indeed, some IPs appeared to change duration by the earlier or later truncation of an underlying inspiratory event. These findings are consistent with the idea that during singing the temporal structure of EPs is under the direct control of forebrain circuits, whereas that of IPs can be strongly influenced by circuits downstream of HVC, likely in the brainstem. An analysis of the temporal jitter of respiratory and vocal structure suggests that IPs may be initiated by HVC at the end of each syllable and terminated by HVC immediately before the onset of the next syllable.
Conflict of interest statement
Figures
Similar articles
-
Temperature Manipulation in Songbird Brain Implicates the Premotor Nucleus HVC in Birdsong Syntax.J Neurosci. 2017 Mar 8;37(10):2600-2611. doi: 10.1523/JNEUROSCI.1827-16.2017. Epub 2017 Feb 3. J Neurosci. 2017. PMID: 28159910 Free PMC article.
-
Learning to breathe and sing: development of respiratory-vocal coordination in young songbirds.J Neurophysiol. 2011 Oct;106(4):1747-65. doi: 10.1152/jn.00247.2011. Epub 2011 Jun 22. J Neurophysiol. 2011. PMID: 21697438 Free PMC article.
-
Telencephalic neurons monosynaptically link brainstem and forebrain premotor networks necessary for song.J Neurosci. 2008 Mar 26;28(13):3479-89. doi: 10.1523/JNEUROSCI.0177-08.2008. J Neurosci. 2008. PMID: 18367614 Free PMC article.
-
Bilateral control and interhemispheric coordination in the avian song motor system.Ann N Y Acad Sci. 2004 Jun;1016:171-86. doi: 10.1196/annals.1298.014. Ann N Y Acad Sci. 2004. PMID: 15313775 Review.
-
Functional organization of forebrain pathways for song production and perception.J Neurobiol. 1997 Nov;33(5):671-93. doi: 10.1002/(sici)1097-4695(19971105)33:5<671::aid-neu12>3.0.co;2-c. J Neurobiol. 1997. PMID: 9369466 Review.
Cited by
-
Model of the songbird nucleus HVC as a network of central pattern generators.J Neurophysiol. 2016 Nov 1;116(5):2405-2419. doi: 10.1152/jn.00438.2016. Epub 2016 Aug 17. J Neurophysiol. 2016. PMID: 27535375 Free PMC article.
-
Predicting plasticity: acute context-dependent changes to vocal performance predict long-term age-dependent changes.J Neurophysiol. 2015 Oct;114(4):2328-39. doi: 10.1152/jn.00688.2015. Epub 2015 Aug 26. J Neurophysiol. 2015. PMID: 26311186 Free PMC article.
-
Thalamus drives vocal onsets in the zebra finch courtship song.Nature. 2023 Apr;616(7955):132-136. doi: 10.1038/s41586-023-05818-x. Epub 2023 Mar 22. Nature. 2023. PMID: 36949189 Free PMC article.
-
Two distinct modes of forebrain circuit dynamics underlie temporal patterning in the vocalizations of young songbirds.J Neurosci. 2011 Nov 9;31(45):16353-68. doi: 10.1523/JNEUROSCI.3009-11.2011. J Neurosci. 2011. PMID: 22072687 Free PMC article.
-
Brain temperature and its fundamental properties: a review for clinical neuroscientists.Front Neurosci. 2014 Oct 8;8:307. doi: 10.3389/fnins.2014.00307. eCollection 2014. Front Neurosci. 2014. PMID: 25339859 Free PMC article. Review.
References
-
- Northcutt RG. Understanding Vertebrate Brain Evolution. Integ and Comp Biol. 2002;42:743–756. - PubMed
-
- Grillner S, Hellgren J, Menard A, Saitoh K, Wikstrom MA. Mechanisms for selection of basic motor programs–roles for the striatum and pallidum. Trends Neurosci. 2005;28:364–370. - PubMed
-
- Moody SL, Zipser D. A model of reaching dynamics in primary motor cortex. J Cognitive Neurosci. 1998;10:35–45. - PubMed
-
- Schwartz AB, Moran DW. Arm trajectory and representation of movement processing in motor cortical activity. Eur J Neurosci. 2000;12:1851–1856. - PubMed
-
- Levelt WJM. Speaking: from intention to articulation: MIT Press 1993
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
