

Changes in early cortical visual processing predict enhanced reactivity in deaf individuals
- PMID: 21980501
- PMCID: PMC3183070
- DOI: 10.1371/journal.pone.0025607
Changes in early cortical visual processing predict enhanced reactivity in deaf individuals
Abstract
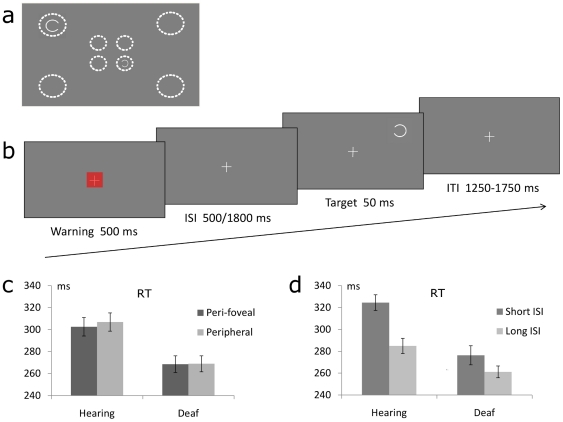
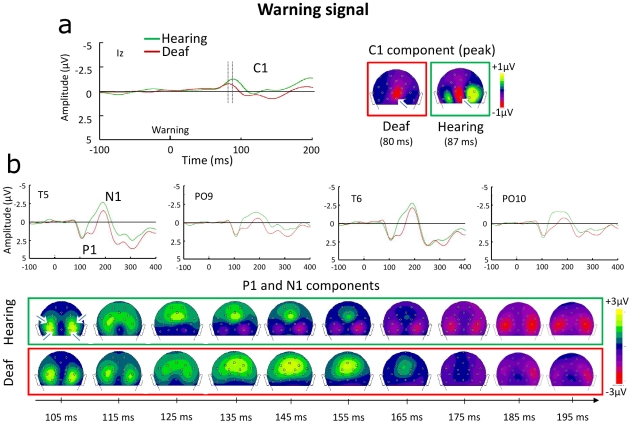
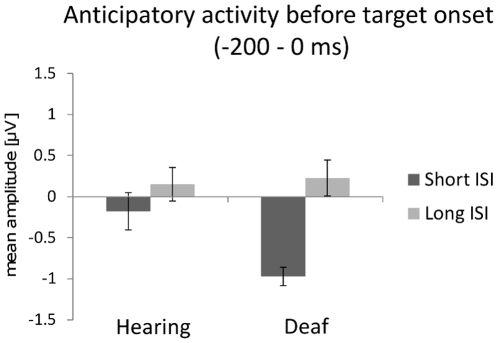
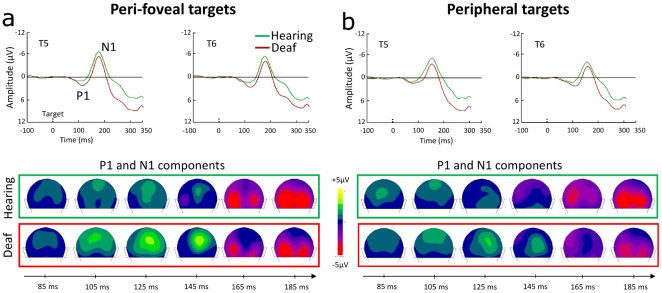
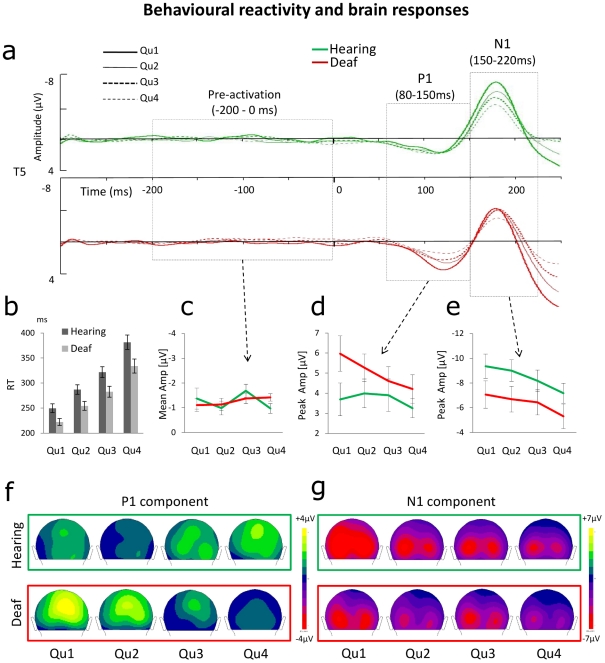
Individuals with profound deafness rely critically on vision to interact with their environment. Improvement of visual performance as a consequence of auditory deprivation is assumed to result from cross-modal changes occurring in late stages of visual processing. Here we measured reaction times and event-related potentials (ERPs) in profoundly deaf adults and hearing controls during a speeded visual detection task, to assess to what extent the enhanced reactivity of deaf individuals could reflect plastic changes in the early cortical processing of the stimulus. We found that deaf subjects were faster than hearing controls at detecting the visual targets, regardless of their location in the visual field (peripheral or peri-foveal). This behavioural facilitation was associated with ERP changes starting from the first detectable response in the striate cortex (C1 component) at about 80 ms after stimulus onset, and in the P1 complex (100-150 ms). In addition, we found that P1 peak amplitudes predicted the response times in deaf subjects, whereas in hearing individuals visual reactivity and ERP amplitudes correlated only at later stages of processing. These findings show that long-term auditory deprivation can profoundly alter visual processing from the earliest cortical stages. Furthermore, our results provide the first evidence of a co-variation between modified brain activity (cortical plasticity) and behavioural enhancement in this sensory-deprived population.
Conflict of interest statement
Figures
References
-
- Loke WH, Song S. Central and peripheral visual processing in hearing and nonhearing individuals. Bulletin of the Psychonomic Society. 1991;29:437–440.
-
- Reynolds HN. Effects of foveal stimulation on peripheral visual processing and laterality in deaf and hearing subjects. The American Journal of Psychology. 1993;106:523–540. - PubMed
-
- Bottari D, Nava E, Ley P, Pavani F. Enhanced reactivity to visual stimuli in deaf individuals. Restorative Neurology and Neuroscience. 2010;28:167–179. - PubMed
-
- Pavani F, Bottari D. Visual abilities in individuals with profound deafness: a critical review. In: Murray MM, Wallace M, editors. Frontiers in the Neural Bases of Multisensory Processes. Boca Raton, FL: CRC Press; in press. - PubMed
-
- Neville HJ, Lawson D. Attention to central and peripheral visual space in a movement detection task: an event-related potential and behavioral study. II. Congenitally deaf adults. Brain Research. 1987;405:268–283. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
