

Dopamine-induced plasticity, phospholipase D (PLD) activity and cocaine-cue behavior depend on PLD-linked metabotropic glutamate receptors in amygdala
- PMID: 21980514
- PMCID: PMC3181343
- DOI: 10.1371/journal.pone.0025639
Dopamine-induced plasticity, phospholipase D (PLD) activity and cocaine-cue behavior depend on PLD-linked metabotropic glutamate receptors in amygdala
Abstract
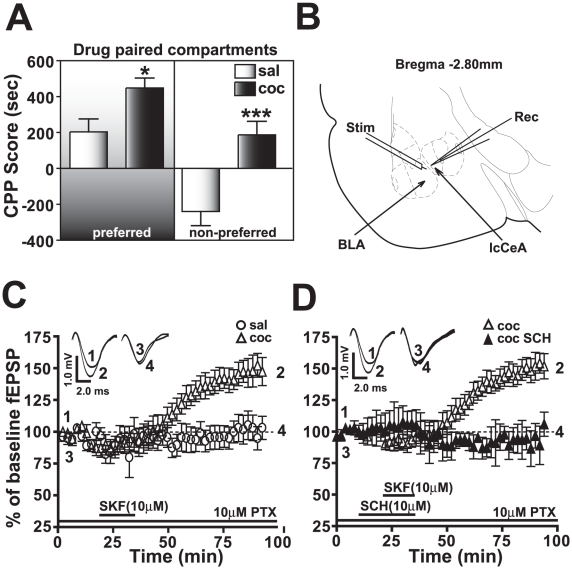
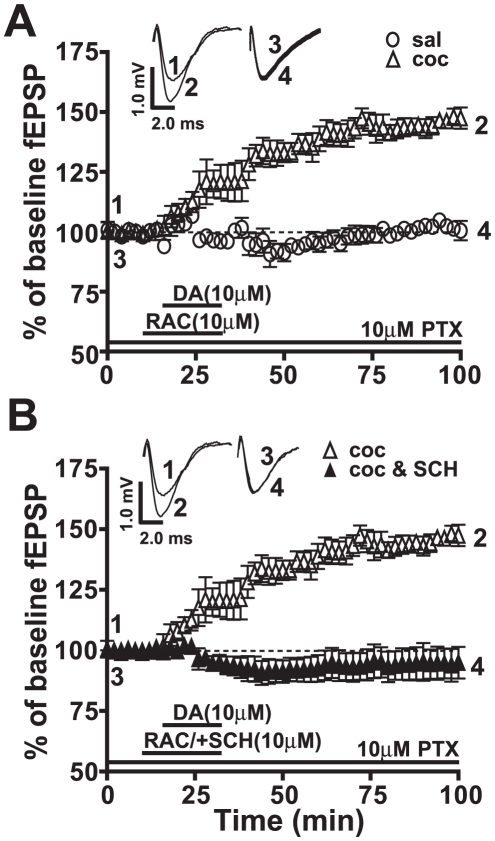
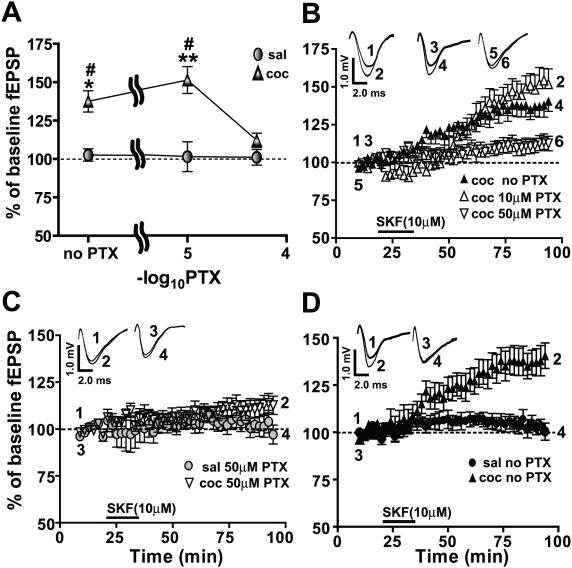
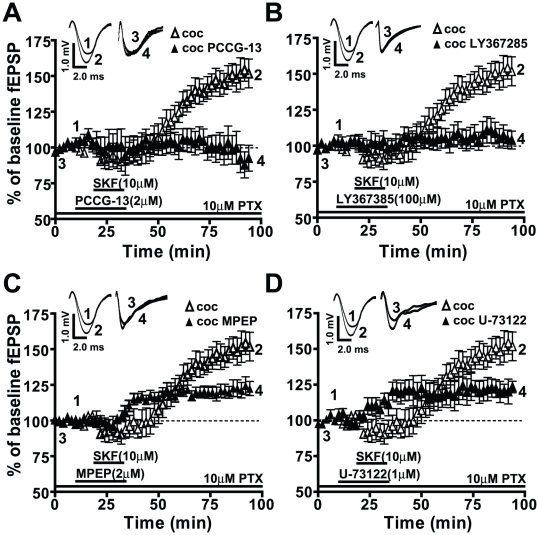
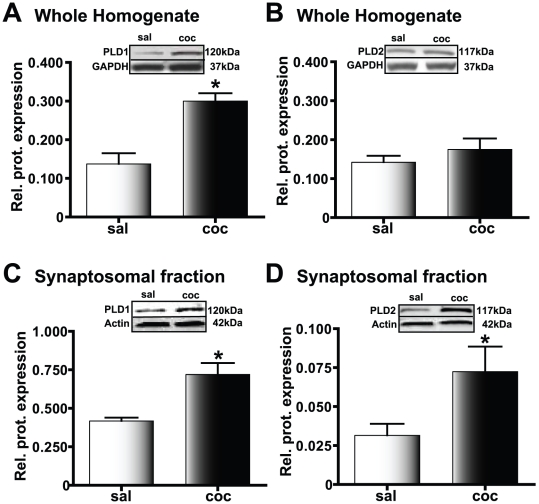
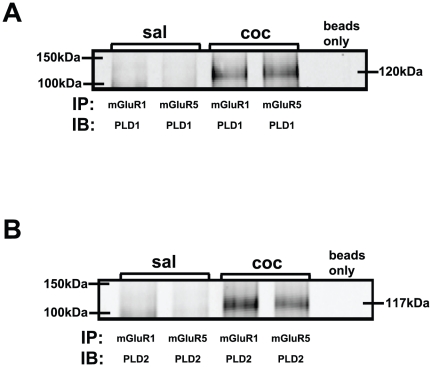
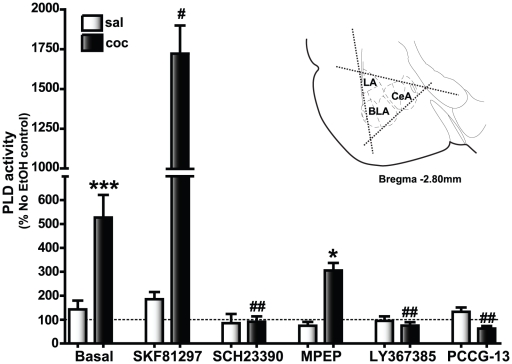
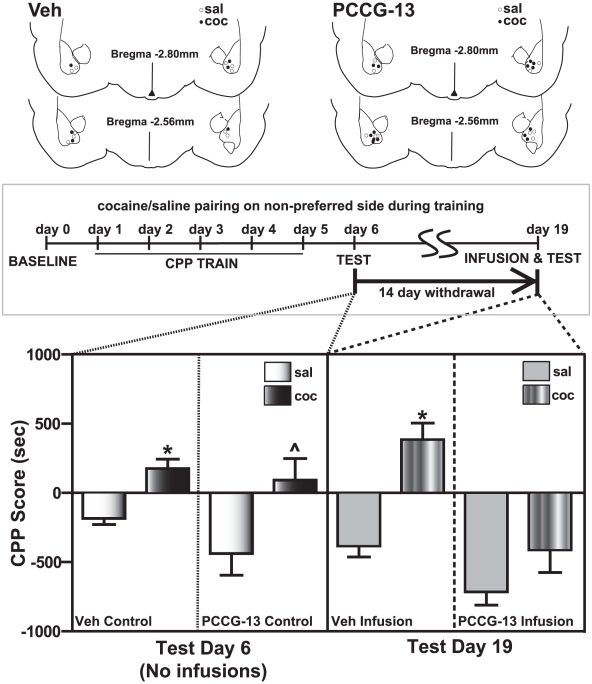
Cocaine-cue associations induce synaptic plasticity with long lasting molecular and cellular changes in the amygdala, a site crucial for cue-associated memory mechanisms. The underlying neuroadaptations can include marked alterations in signaling via dopamine (DA) receptors (DRs) and metabotropic glutamate (Glu) receptors (mGluRs). Previously, we reported that DR antagonists blocked forms of synaptic plasticity in amygdala slices of Sprague-Dawley rats withdrawn from repeated cocaine administration. In the present study, we investigated synaptic plasticity induced by exogenous DA and its dependence on mGluR signaling and a potential role for phospholipase D (PLD) as a downstream element linked to mGluR and DR signaling. Utilizing a modified conditioned place preference (CPP) paradigm as a functional behavioral measure, we studied the neurophysiological effects after two-weeks to the last cocaine conditioning. We recorded, electrophysiologically, a DR-induced synaptic potentiation in the basolateral to lateral capsula central amygdala (BLA-lcCeA) synaptic pathway that was blocked by antagonists of group I mGluRs, particularly, the PLD-linked mGluR. In addition, we observed 2-2.5 fold increase in PLD expression and 3.7-fold increase in basal PLD enzyme activity. The enhanced PLD activity could be further stimulated (9.3 fold) by a DA D1-like (D1/5R) receptor agonist, and decreased to control levels by mGluR1 and PLD-linked mGluR antagonists. Diminished CPP was observed by infusion of a PLD-linked mGluR antagonist, PCCG-13, in the amygdala 15 minutes prior to testing, two weeks after the last cocaine injection. These results imply a functional interaction between D1/5Rs, group I mGluRs via PLD in the amygdala synaptic plasticity associated with cocaine-cues.
Conflict of interest statement
Figures
Similar articles
-
Fear potentiated startle increases phospholipase D (PLD) expression/activity and PLD-linked metabotropic glutamate receptor mediated post-tetanic potentiation in rat amygdala.Neurobiol Learn Mem. 2016 Feb;128:65-79. doi: 10.1016/j.nlm.2015.12.009. Epub 2015 Dec 31. Neurobiol Learn Mem. 2016. PMID: 26748024 Free PMC article.
-
Cocaine withdrawal reduces group I mGluR-mediated long-term potentiation via decreased GABAergic transmission in the amygdala.Eur J Neurosci. 2011 Jul;34(2):177-89. doi: 10.1111/j.1460-9568.2011.07769.x. Epub 2011 Jul 12. Eur J Neurosci. 2011. PMID: 21749491 Free PMC article.
-
Cocaine and kindling alter the sensitivity of group II and III metabotropic glutamate receptors in the central amygdala.J Neurophysiol. 2000 Aug;84(2):759-70. doi: 10.1152/jn.2000.84.2.759. J Neurophysiol. 2000. PMID: 10938303
-
Using metabotropic glutamate receptors to modulate cocaine's synaptic and behavioral effects: mGluR1 finds a niche.Curr Opin Neurobiol. 2013 Aug;23(4):500-6. doi: 10.1016/j.conb.2013.01.009. Epub 2013 Feb 4. Curr Opin Neurobiol. 2013. PMID: 23385114 Free PMC article. Review.
-
Dopamine D1/D5 receptors mediate informational saliency that promotes persistent hippocampal long-term plasticity.Cereb Cortex. 2014 Apr;24(4):845-58. doi: 10.1093/cercor/bhs362. Epub 2012 Nov 25. Cereb Cortex. 2014. PMID: 23183712 Free PMC article. Review.
Cited by
-
Inhibitory effects of dopamine agonists on pain-responsive neurons in the central nucleus of the amygdala.J Neurophysiol. 2025 Jun 1;133(6):1947-1954. doi: 10.1152/jn.00135.2025. Epub 2025 May 27. J Neurophysiol. 2025. PMID: 40423681 Free PMC article.
-
Inhibition of phospholipase D promotes neurological function recovery and reduces neuroinflammation after spinal cord injury in mice.Front Cell Neurosci. 2024 Mar 20;18:1352630. doi: 10.3389/fncel.2024.1352630. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38572075 Free PMC article.
-
Corticotropin releasing factor and catecholamines enhance glutamatergic neurotransmission in the lateral subdivision of the central amygdala.Neuropharmacology. 2013 Jul;70:316-23. doi: 10.1016/j.neuropharm.2013.02.014. Epub 2013 Mar 5. Neuropharmacology. 2013. PMID: 23470280 Free PMC article.
-
Fear potentiated startle increases phospholipase D (PLD) expression/activity and PLD-linked metabotropic glutamate receptor mediated post-tetanic potentiation in rat amygdala.Neurobiol Learn Mem. 2016 Feb;128:65-79. doi: 10.1016/j.nlm.2015.12.009. Epub 2015 Dec 31. Neurobiol Learn Mem. 2016. PMID: 26748024 Free PMC article.
-
Amygdala-Hippocampal Phospholipase D (PLD) Signaling As Novel Mechanism of Cocaine-Environment Maladaptive Conditioned Responses.Int J Neuropsychopharmacol. 2016 Jun;19(6):pyv139. doi: 10.1093/ijnp/pyv139. Epub 2016 Jan 22. Int J Neuropsychopharmacol. 2016. PMID: 26802567 Free PMC article.
References
-
- Hyman SE. Addiction: a disease of learning and memory. Am J Psychiatry. 2005;162:1414–1422. - PubMed
-
- Buffalari DM, See RE. Amygdala mechanisms of Pavlovian psychostimulant conditioning and relapse. Curr Top Behav Neurosci. 2010;3:73–99. - PubMed
-
- Hyman SE, Malenka RC, Nestler EJ. Neural mechanisms of addiction: the role of reward-related learning and memory. Annu Rev Neurosci. 2006;29:565–598. - PubMed
-
- Breiter HC, Gollub RL, Weisskoff RM, Kennedy DN, Makris N, et al. Acute effects of cocaine on human brain activity and emotion. Neuron. 1997;19:591–611. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
