

Cohesin's concatenation of sister DNAs maintains their intertwining
- PMID: 21981921
- PMCID: PMC3240746
- DOI: 10.1016/j.molcel.2011.07.034
Cohesin's concatenation of sister DNAs maintains their intertwining
Abstract
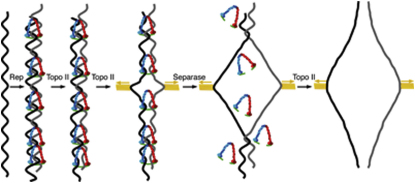
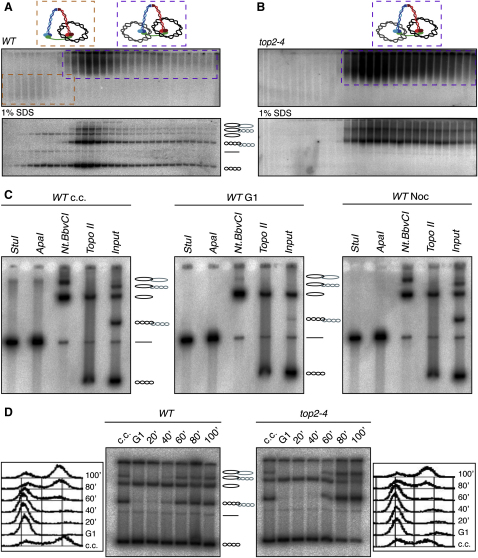
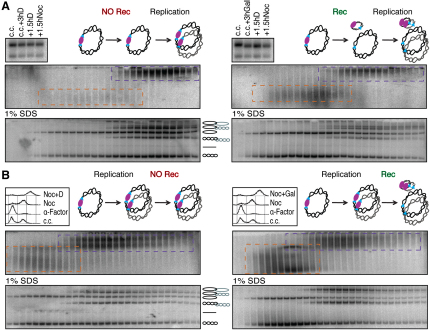
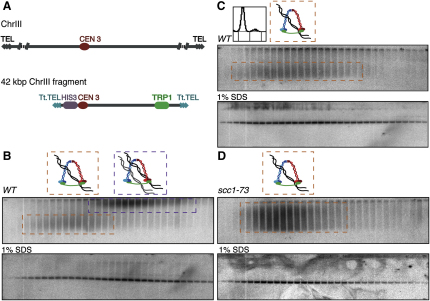
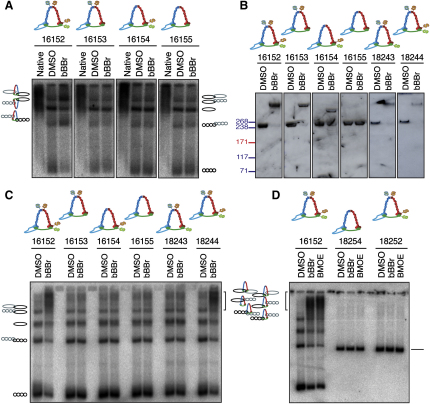
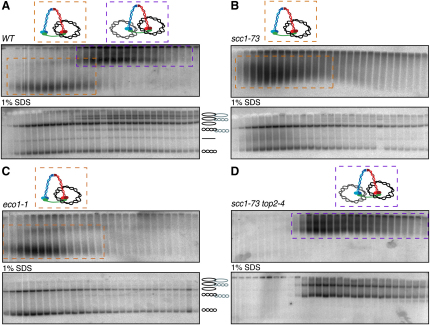
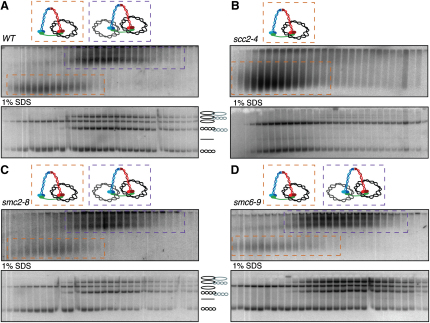
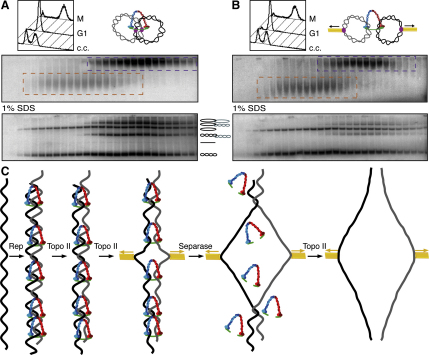
The contribution of DNA catenation to sister chromatid cohesion is unclear partly because it has never been observed directly within mitotic chromosomes. Differential sedimentation-velocity and gel electrophoresis reveal that sisters of 26 kb circular minichromosomes are held together by catenation as well as by cohesin. The finding that chemical crosslinking of cohesin's three subunit interfaces entraps sister DNAs of circular but not linear minichromosomes implies that cohesin functions using a topological principle. Importantly, cohesin holds both catenated and uncatenated DNAs together in this manner. In the vicinity of centromeres, catenanes are resolved by spindle forces, but linkages mediated directly by cohesin resist these forces even after complete decatenation. Crucially, persistence of catenation after S phase depends on cohesin. We conclude that by retarding Topo II-driven decatenation, cohesin mediates sister chromatid cohesion by an indirect mechanism as well as one involving entrapment of sister DNAs inside its tripartite ring.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A double lock on sister chromatids by cohesin.Mol Cell. 2011 Oct 7;44(1):5-6. doi: 10.1016/j.molcel.2011.09.008. Mol Cell. 2011. PMID: 21981913
References
-
- Baxter J., Sen N., Martínez V.L., De Carandini M.E.M., Schvartzman J.B., Diffley J.F.X., Aragón L. Positive supercoiling of mitotic DNA drives decatenation by topoisomerase II in eukaryotes. Science. 2011;331:1328–1332. - PubMed
-
- Ciosk R., Shirayama M., Shevchenko A., Tanaka T., Toth A., Shevchenko A., Nasmyth K. Cohesin's binding to chromosomes depends on a separate complex consisting of Scc2 and Scc4 proteins. Mol. Cell. 2000;5:243–254. - PubMed
-
- Coelho P.A., Queiroz-Machado J., Sunkel C.E. Condensin-dependent localisation of topoisomerase II to an axial chromosomal structure is required for sister chromatid resolution during mitosis. J. Cell Sci. 2003;116:4763–4776. - PubMed
-
- D'Ambrosio C., Kelly G., Shirahige K., Uhlmann F. Condensin-dependent rDNA decatenation introduces a temporal pattern to chromosome segregation. Curr. Biol. 2008;18:1084–1089. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
