

Acetylation regulates the stability of a bacterial protein: growth stage-dependent modification of RNase R
- PMID: 21981926
- PMCID: PMC3191462
- DOI: 10.1016/j.molcel.2011.06.037
Acetylation regulates the stability of a bacterial protein: growth stage-dependent modification of RNase R
Abstract
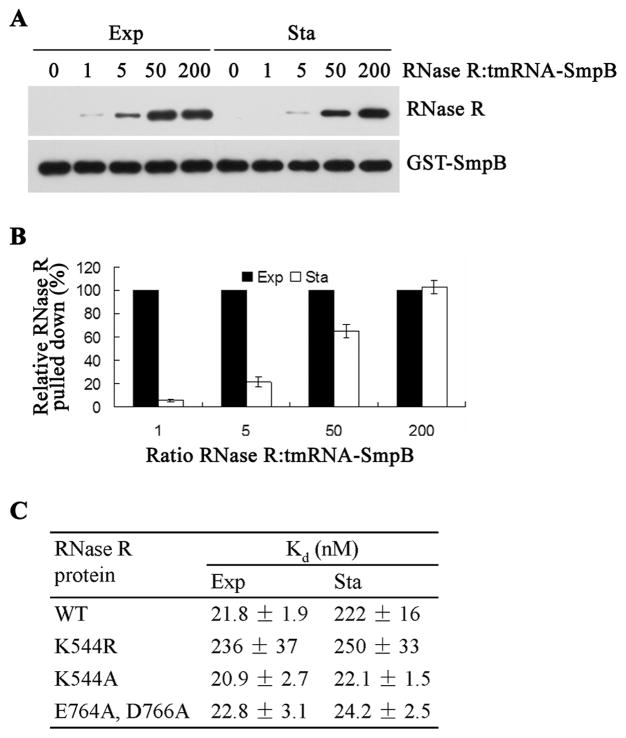
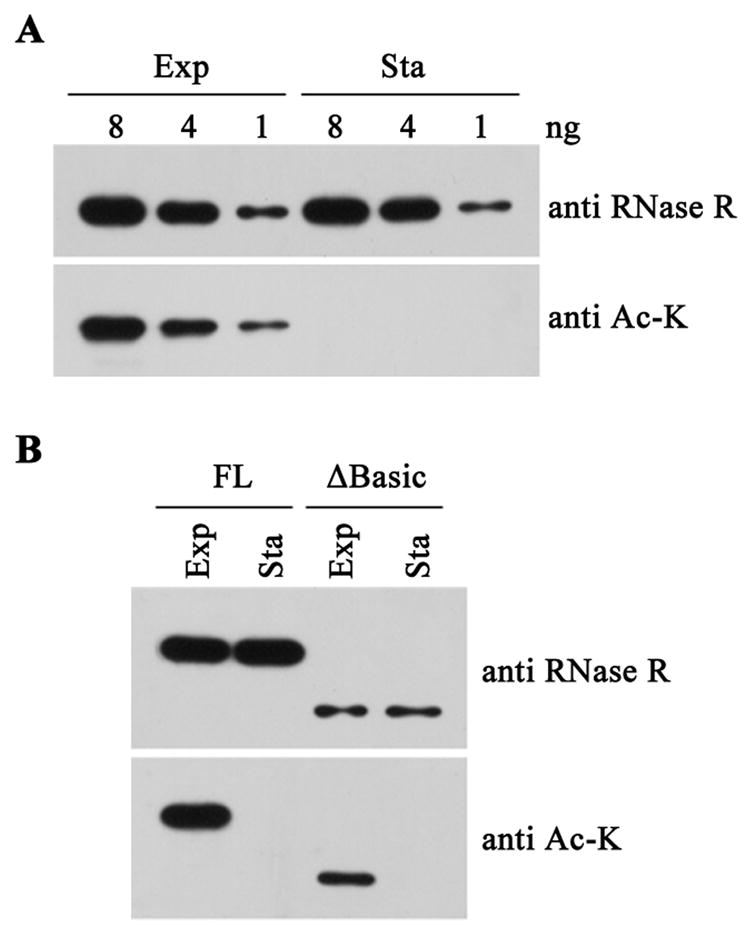
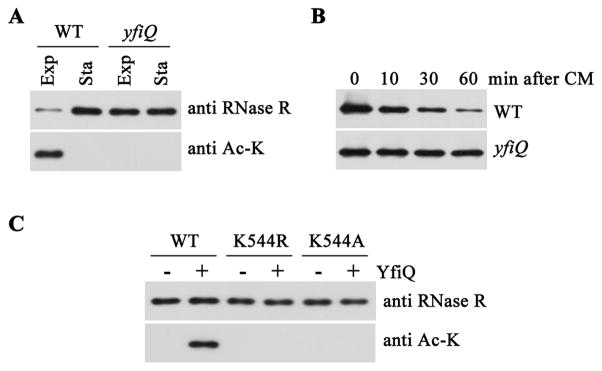
RNase R, an Escherichia coli exoribonuclease important for degradation of structured RNAs, increases 3- to 10-fold under certain stress conditions, due to an increased half-life for this usually unstable protein. Components of the trans-translation machinery, tmRNA, and its associated protein, SmpB, are essential for RNase R instability. However, it is not understood why exponential phase RNase R is unstable or how it becomes stabilized in stationary phase. Here, we show that these phenomena are regulated by acetylation catalyzed by YfiQ protein. One residue, Lys544, is acetylated in exponential phase RNase R, but not in the stationary phase protein, resulting in tighter binding of tmRNA-SmpB to the C-terminal region of exponential phase RNase R and subsequent proteolytic degradation. Removal of the positive charge at Lys544 or a negative charge in the C-terminal region likely disrupts their interaction, facilitating tmRNA-SmpB binding. These findings indicate that acetylation can regulate the stability of a bacterial protein.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures

References
-
- Andrade JM, Cairrão F, Arraiano CM. RNase R affects gene expression in stationary phase: Regulation of ompA. Mol Microbiol. 2006;60:219–228. - PubMed
-
- Andrade JM, Pobre V, Silva IJ, Domingues S, Arraiano CM. The role of 3′-5′ exoribonucleases in RNA degradation. Prog Mol Biol Transl Sci. 2009;85:187–229. - PubMed
-
- Blander G, Guarente L. The Sir2 family of protein deacetylases. Annu Rev Biochem. 2004;73:417–435. - PubMed
-
- Bordone L, Guarente L. Calorie restriction, SIRT1 and metabolism: understanding longevity. Nat Rev Mol Cell Biol. 2005;6:298–305. - PubMed
-
- Cairrão F, Cruz A, Mori H, Arraiano CM. Cold shock induction of RNase R and its role in the maturation of the quality control mediator SsrA/tmRNA. Mol Microbiol. 2003;50:1349–1360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
