

Two forms of loops generate the chromatin conformation of the immunoglobulin heavy-chain gene locus
- PMID: 21982154
- PMCID: PMC3685183
- DOI: 10.1016/j.cell.2011.08.049
Two forms of loops generate the chromatin conformation of the immunoglobulin heavy-chain gene locus
Abstract
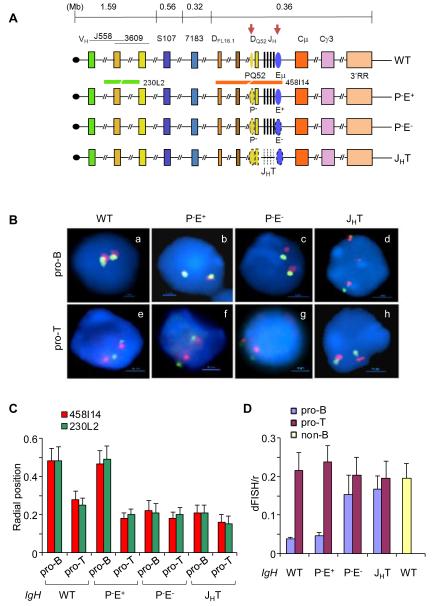
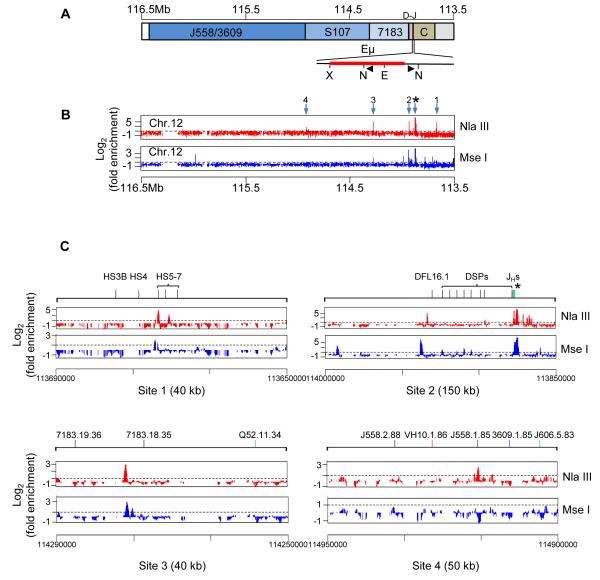
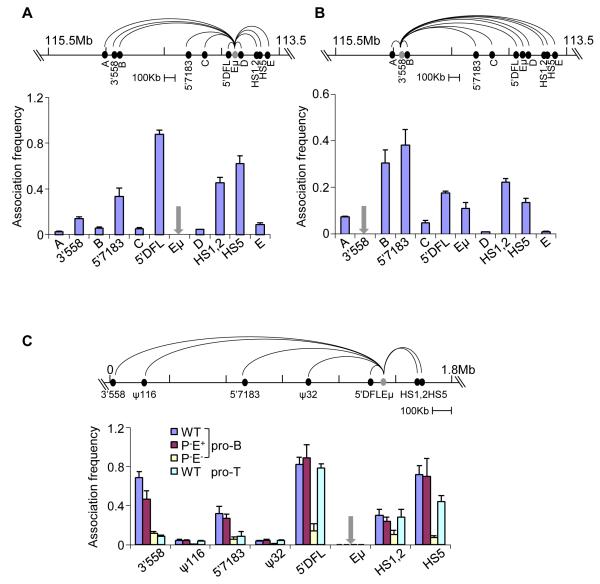
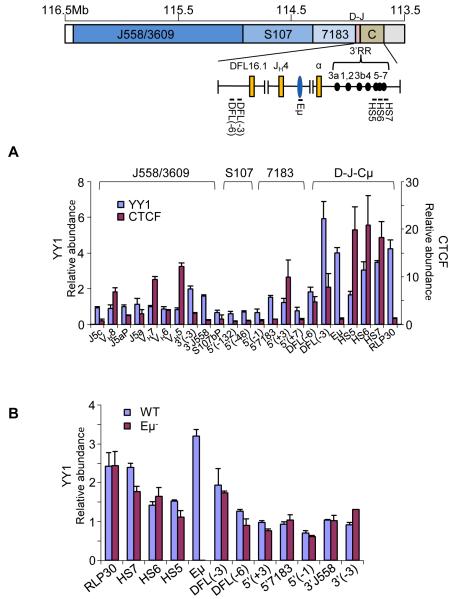
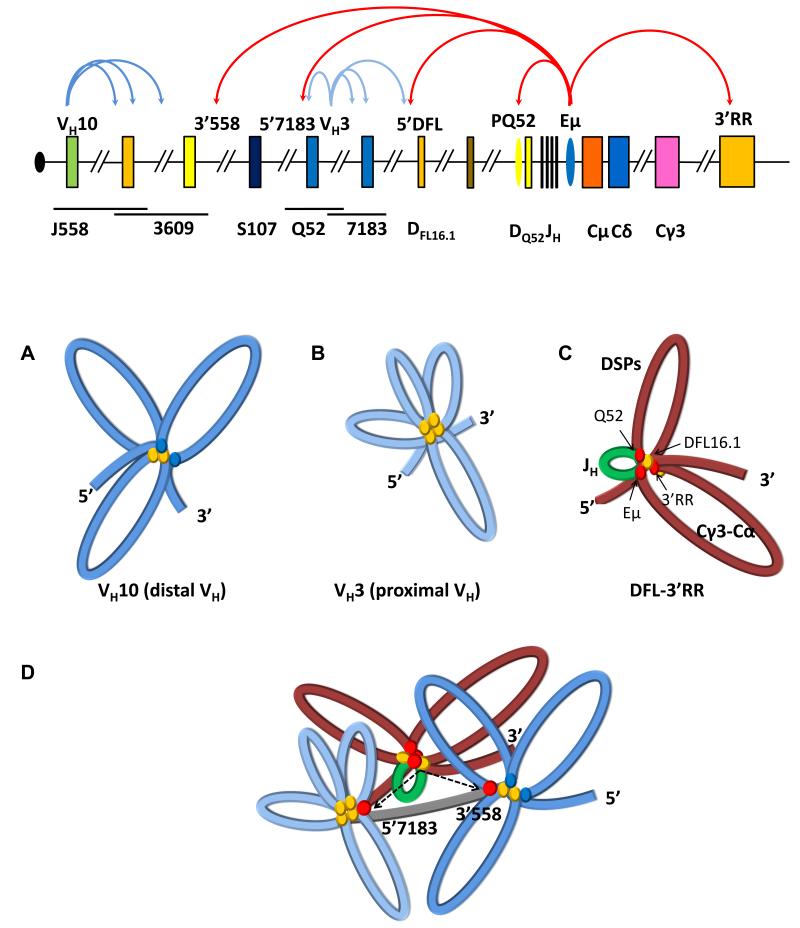
The immunoglobulin heavy-chain (IgH) gene locus undergoes radial repositioning within the nucleus and locus contraction in preparation for gene recombination. We demonstrate that IgH locus conformation involves two levels of chromosomal compaction. At the first level, the locus folds into several multilooped domains. One such domain at the 3' end of the locus requires an enhancer, Eμ; two other domains at the 5' end are Eμ independent. At the second level, these domains are brought into spatial proximity by Eμ-dependent interactions with specific sites within the V(H) region. Eμ is also required for radial repositioning of IgH alleles, indicating its essential role in large-scale chromosomal movements in developing lymphocytes. Our observations provide a comprehensive view of the conformation of IgH alleles in pro-B cells and the mechanisms by which it is established.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


References
-
- Afshar R, Pierce S, Bolland DJ, Corcoran A, Oltz EM. Regulation of IgH gene assembly: role of the intronic enhancer and 5′DQ52 region in targeting DHJH recombination. J Immunol. 2006;176:2439–2447. - PubMed
-
- Chakraborty T, Chowdhury D, Keyes A, Jani A, Subrahmanyam R, Ivanova I, Sen R. Repeat organization and epigenetic regulation of the DH-Cmu domain of the immunoglobulin heavy-chain gene locus. Mol Cell. 2007;27:842–850. - PubMed
-
- Cook PR, Brazell IA, Jost E. Characterization of nuclear structures containing superhelical DNA. J Cell Sci. 1976;22:303–324. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
