

The tubulin deglutamylase CCPP-1 regulates the function and stability of sensory cilia in C. elegans
- PMID: 21982591
- PMCID: PMC4680987
- DOI: 10.1016/j.cub.2011.08.049
The tubulin deglutamylase CCPP-1 regulates the function and stability of sensory cilia in C. elegans
Abstract
Background: Posttranslational modifications (PTMs) such as acetylation, detyrosination, and polyglutamylation have long been considered markers of stable microtubules and have recently been proposed to guide molecular motors to specific subcellular destinations. Microtubules can be deglutamylated by the cytosolic carboxypeptidase CCP1. Loss of CCP1 in mice causes cerebellar Purkinje cell degeneration. Cilia, which are conserved organelles that play important diverse roles in animal development and sensation, contain axonemes comprising microtubules that are especially prone to PTMs.
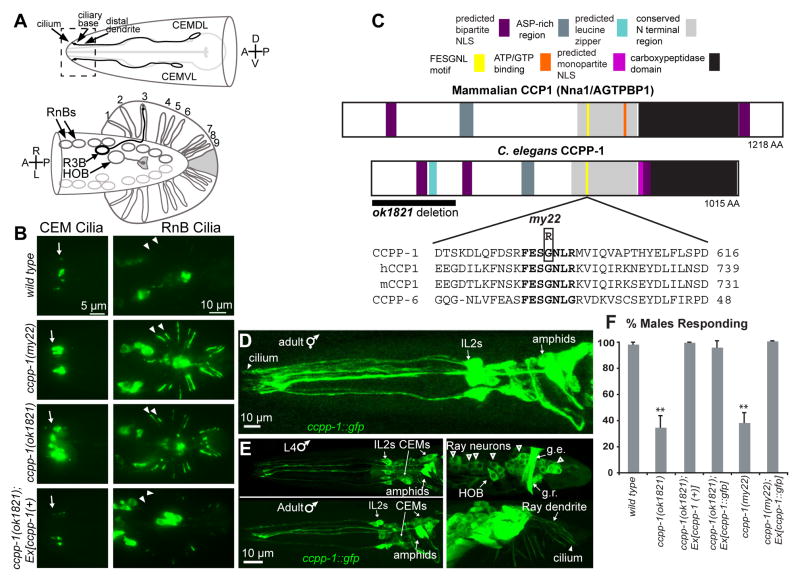
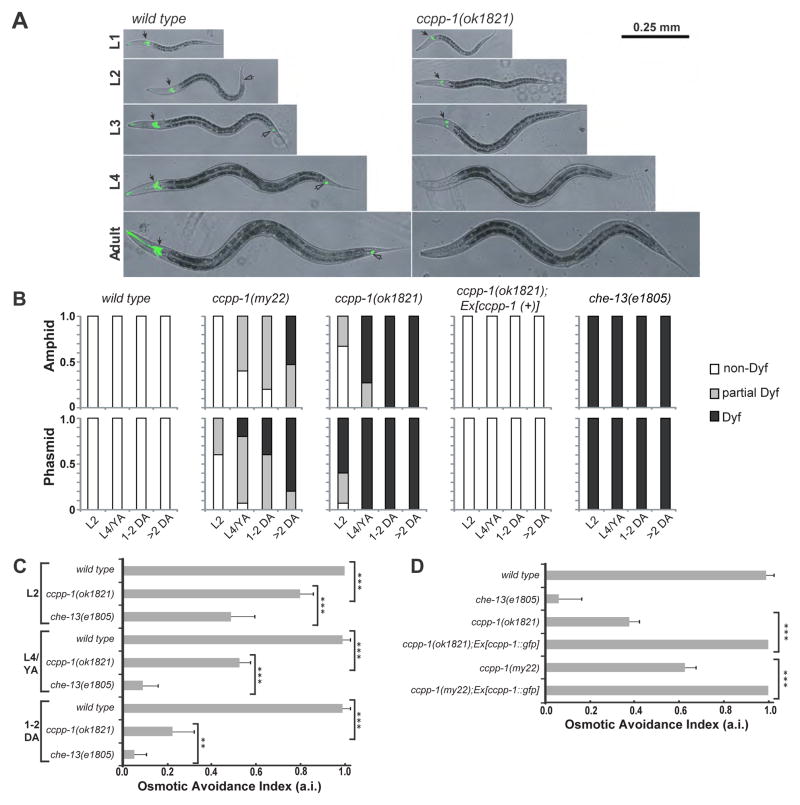
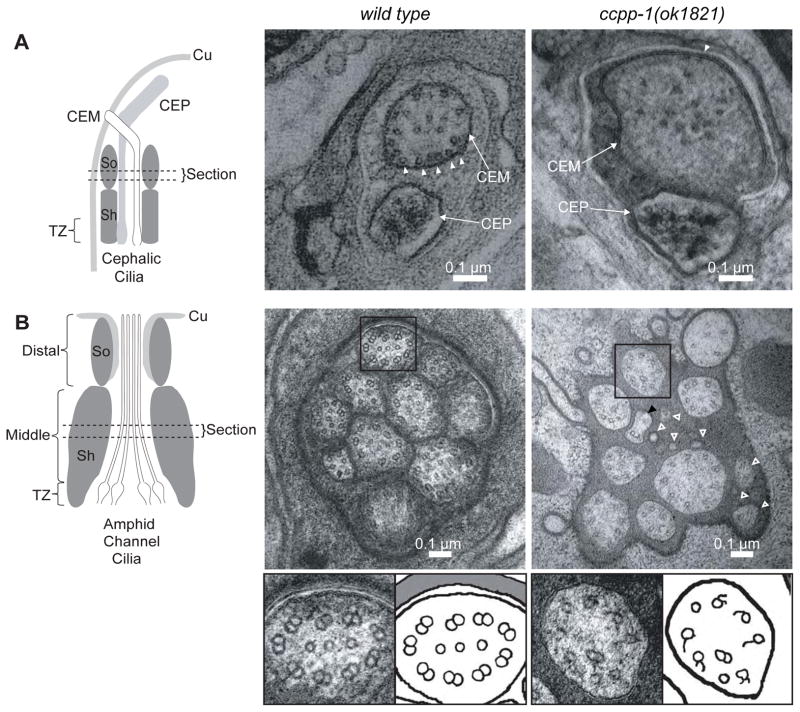
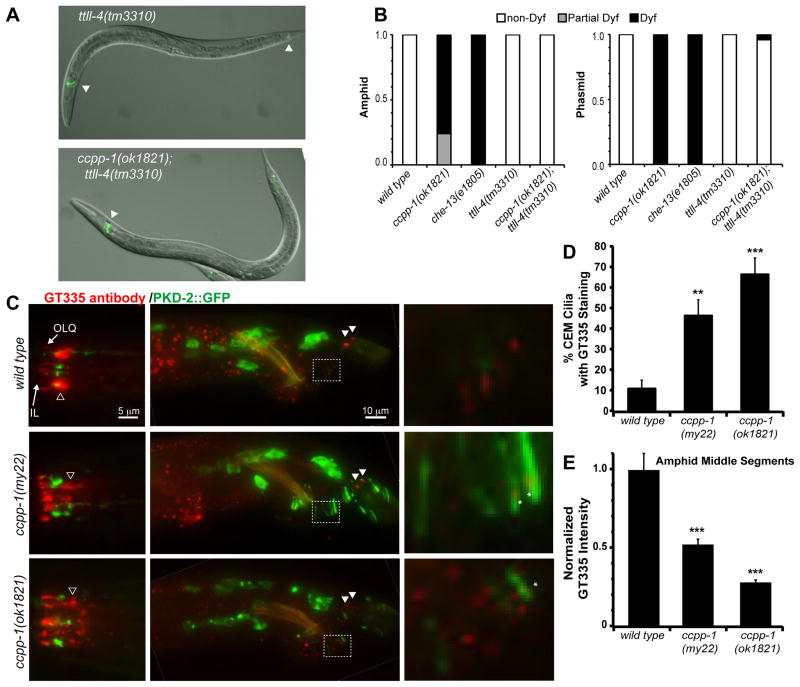
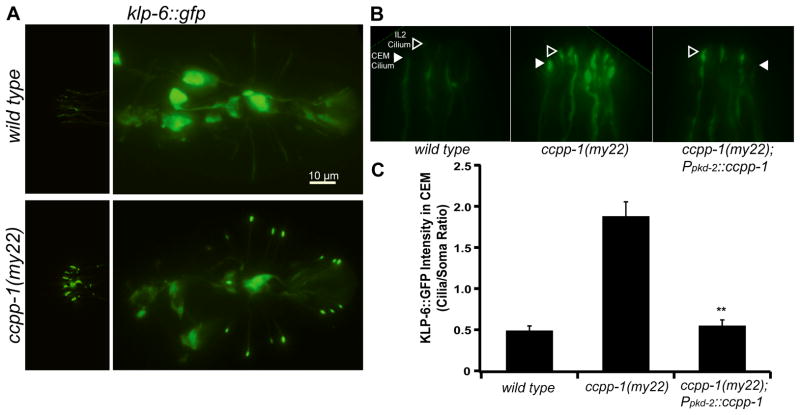
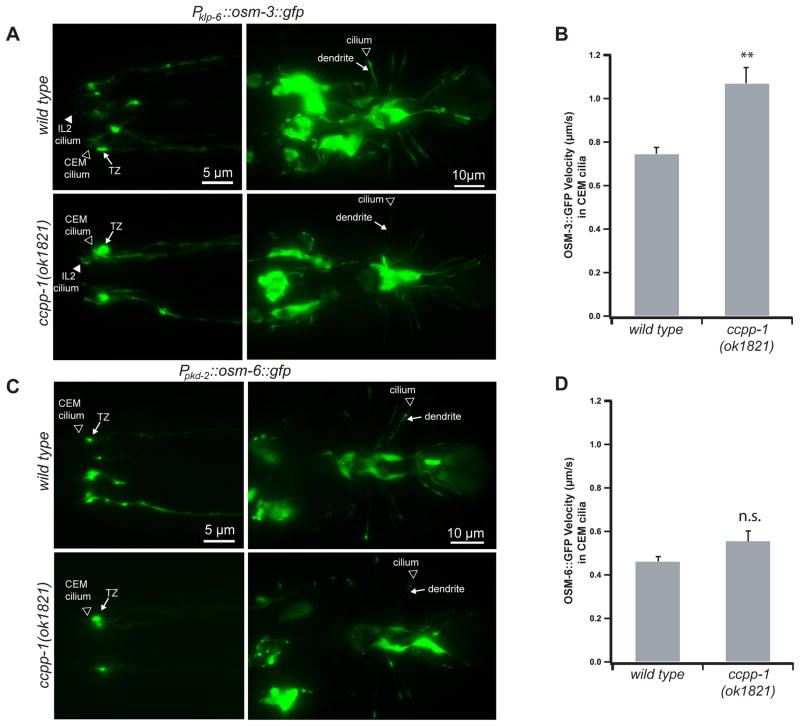
Results: Here, we report that a CCP1 homolog, CCPP-1, regulates the ciliary localization of the kinesin-3 KLP-6 and the polycystin PKD-2 in male-specific sensory neurons in C. elegans. In male-specific CEM (cephalic sensilla, male) cilia, ccpp-1 also controls the velocity of the kinesin-2 OSM-3/KIF17 without affecting the transport of kinesin-II cargo. In the core ciliated nervous system of both males and hermaphrodites, loss of ccpp-1 causes progressive defects in amphid and phasmid sensory cilia, suggesting that CCPP-1 activity is required for ciliary maintenance but not ciliogenesis. Affected cilia exhibit defective B-tubules. Loss of TTLL-4, a polyglutamylating enzyme of the tubulin tyrosine ligase-like family, suppresses progressive ciliary defects in ccpp-1 mutants.
Conclusions: Our studies suggest that CCPP-1 acts as a tubulin deglutamylase that regulates the localization and velocity of kinesin motors and the structural integrity of microtubules in sensory cilia of a multicellular, living animal. We propose that the neuronal degeneration caused by loss of CCP1 in mammals may represent a novel ciliopathy in which cilia are formed but not maintained, depriving the cell of cilia-based signal transduction.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Fliegauf M, Benzing T, Omran H. When cilia go bad: cilia defects and ciliopathies. Nat Rev Mol Cell Biol. 2007;8:880–893. - PubMed
-
- Bulinski JC, Gundersen GG. Stabilization of post-translational modification of microtubules during cellular morphogenesis. BioEssays : news and reviews in molecular, cellular and developmental biology. 1991;13:285–293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
