

Differential state-dependent modification of rat Na(v)1.6 sodium channels expressed in human embryonic kidney (HEK293) cells by the pyrethroid insecticides tefluthrin and deltamethrin
- PMID: 21983428
- PMCID: PMC3226847
- DOI: 10.1016/j.taap.2011.09.021
Differential state-dependent modification of rat Na(v)1.6 sodium channels expressed in human embryonic kidney (HEK293) cells by the pyrethroid insecticides tefluthrin and deltamethrin
Abstract
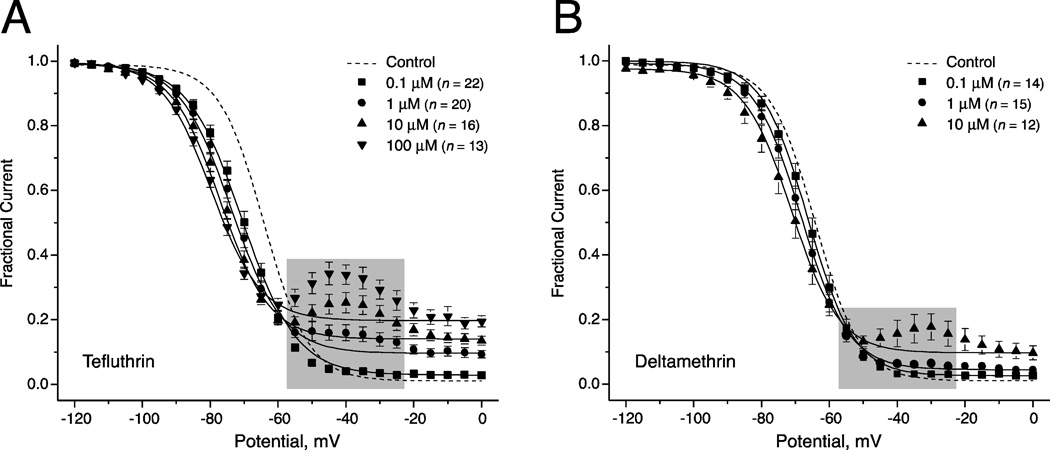
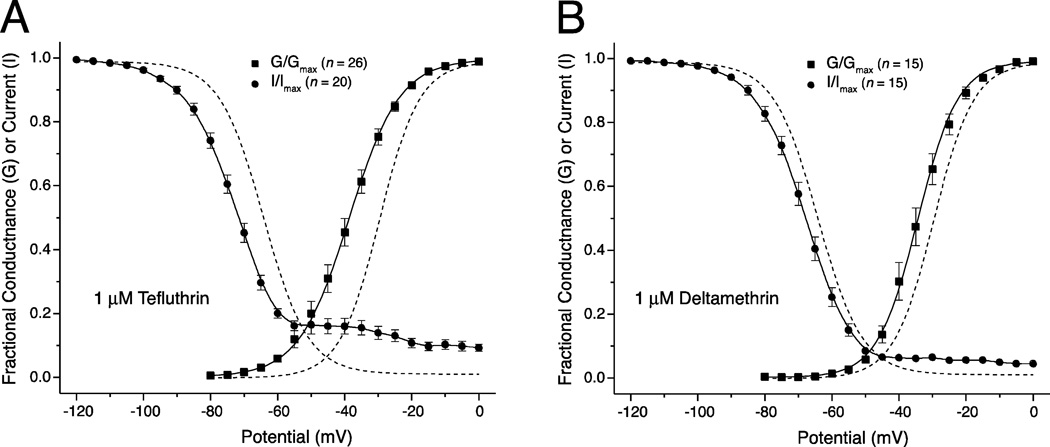
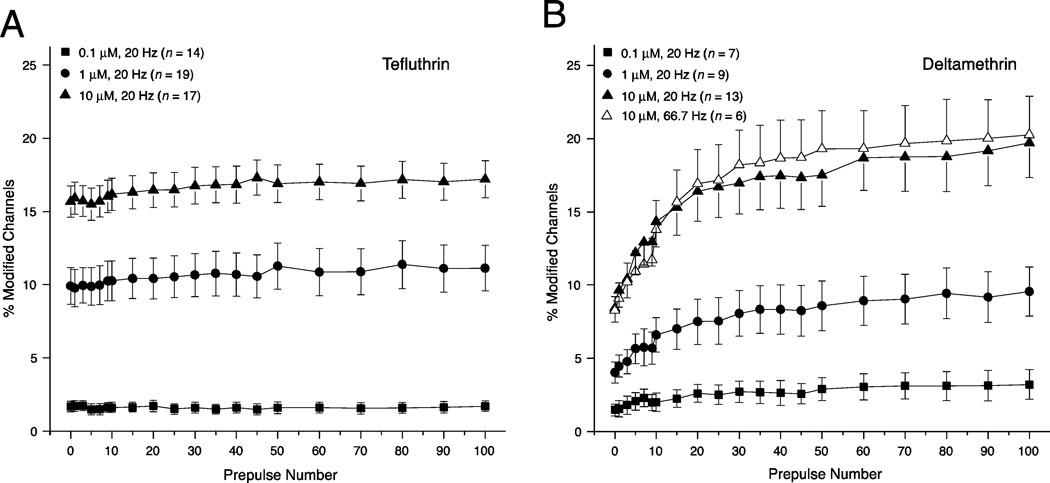
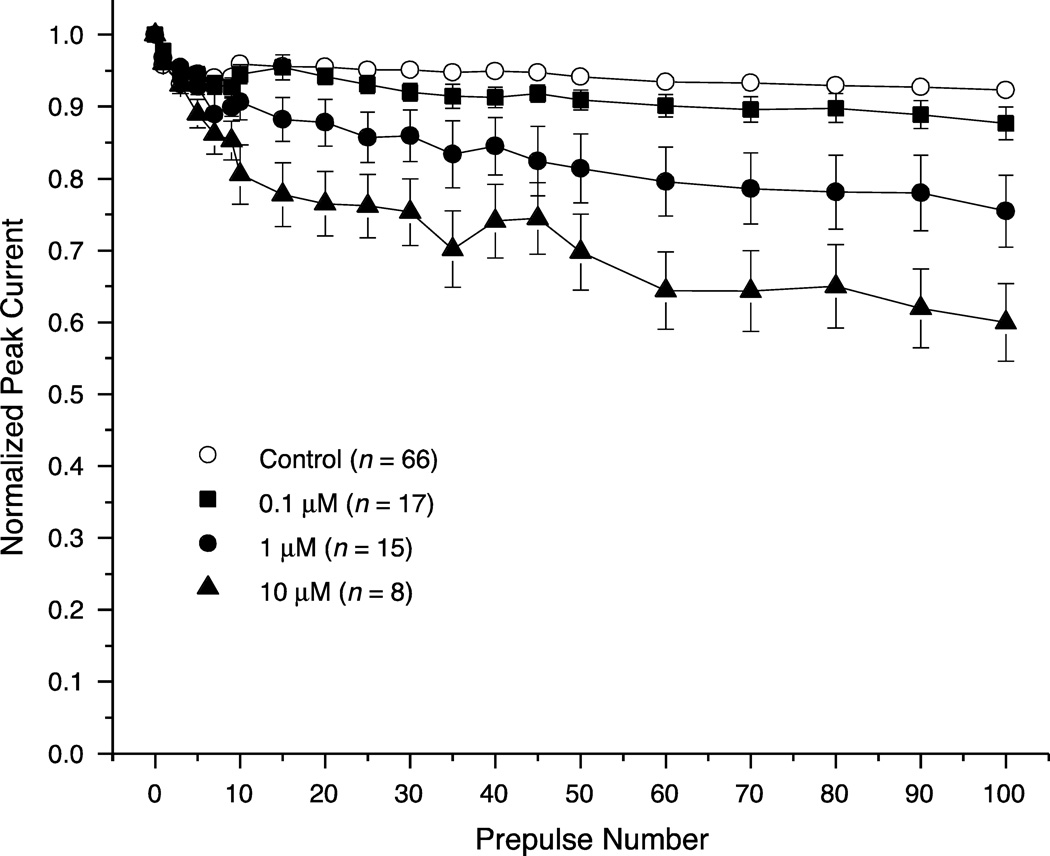
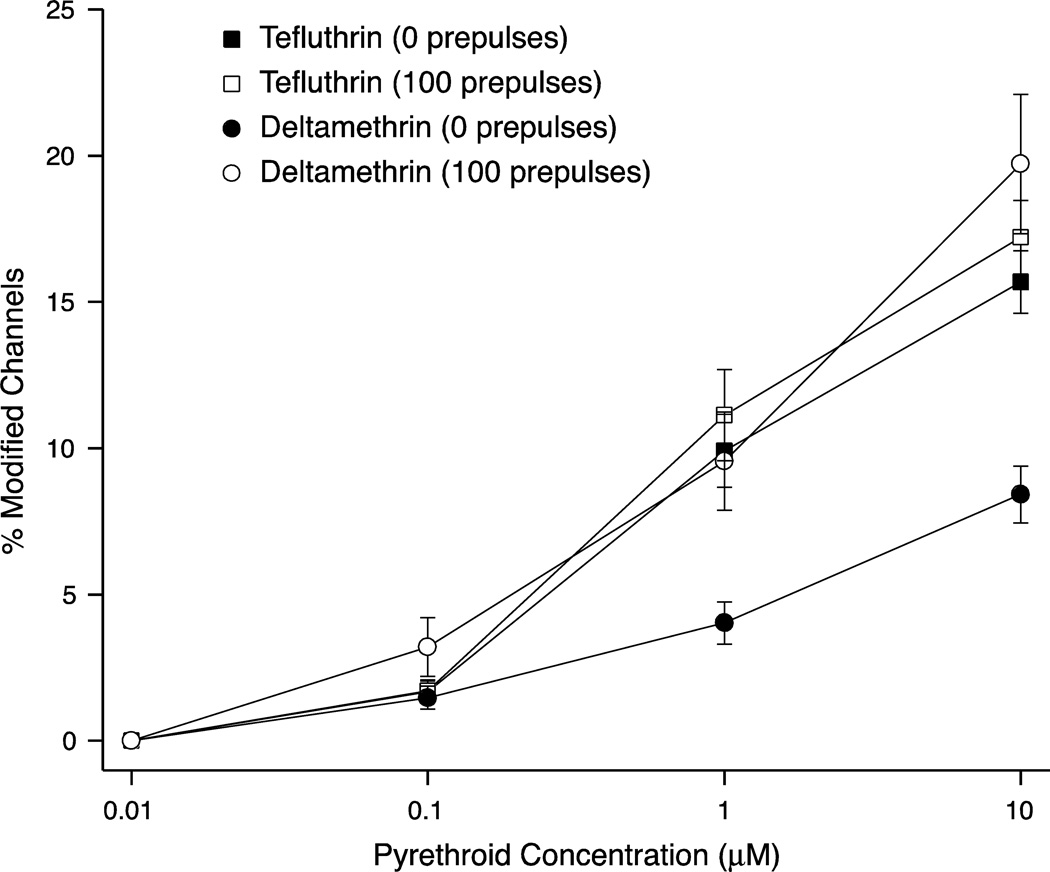
We expressed rat Na(v)1.6 sodium channels in combination with the rat β1 and β2 auxiliary subunits in human embryonic kidney (HEK293) cells and evaluated the effects of the pyrethroid insecticides tefluthrin and deltamethrin on expressed sodium currents using the whole-cell patch clamp technique. Both pyrethroids produced concentration-dependent, resting modification of Na(v)1.6 channels, prolonging the kinetics of channel inactivation and deactivation to produce persistent "late" currents during depolarization and tail currents following repolarization. Both pyrethroids also produced concentration dependent hyperpolarizing shifts in the voltage dependence of channel activation and steady-state inactivation. Maximal shifts in activation, determined from the voltage dependence of the pyrethroid-induced late and tail currents, were ~25mV for tefluthrin and ~20mV for deltamethrin. The highest attainable concentrations of these compounds also caused shifts of ~5-10mV in the voltage dependence of steady-state inactivation. In addition to their effects on the voltage dependence of inactivation, both compounds caused concentration-dependent increases in the fraction of sodium current that was resistant to inactivation following strong depolarizing prepulses. We assessed the use-dependent effects of tefluthrin and deltamethrin on Na(v)1.6 channels by determining the effect of trains of 1 to 100 5-ms depolarizing prepulses at frequencies of 20 or 66.7Hz on the extent of channel modification. Repetitive depolarization at either frequency increased modification by deltamethrin by ~2.3-fold but had no effect on modification by tefluthrin. Tefluthrin and deltamethrin were equally potent as modifiers of Na(v)1.6 channels in HEK293 cells using the conditions producing maximal modification as the basis for comparison. These findings show that the actions of tefluthrin and deltamethrin of Na(v)1.6 channels in HEK293 cells differ from the effects of these compounds on Na(v)1.6 channels in Xenopus oocytes and more closely reflect the actions of pyrethroids on channels in their native neuronal environment.
Copyright © 2011 Elsevier Inc. All rights reserved.
Conflict of interest statement
Neither B. He nor D. M. Soderlund have conflicts of interest regarding the research described in this manuscript.
Figures
References
-
- Ahn H-S, Dib-Hajj SD, Cox JJ, Tyrrell L, Elmslie FV, Clarke AA, Drenth JPH, Woods CG, Waxman SG. A new sodium channel gene mutation I234T in a child with severe pain. European Journal of Pain. 2010;14:944–950. - PubMed
-
- Attwell D, Cohen I, Eisner D, Ohba M, Ojeda C. The steady state TTX-sensitive ("window") sodium current in cardiac Purkinje fibres. Pflugers Arch. 1979;379:137–142. - PubMed
-
- Auld VJ, Goldin AL, Krafte DS, Marshall J, Dunn JM, Catterall WA, Lester HA, Davidson N, Dunn RJ. A rat brain Na+ channel α subunit with novel gating properties. Neuron. 1988;1:449–461. - PubMed
-
- Breckenridge CB, Holden L, Sturgess N, Weiner M, Sheets L, Sargent D, Soderlund DM, Choi J-S, Symington S, Clark JM, Burr S, Ray D. Evidence for a separate mechanism of toxicity for the Type I and Type II pyrethroid insecticides. Neurotoxicology. 2009;30:S17–S31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
