

Microtubules induce self-organization of polarized PAR domains in Caenorhabditis elegans zygotes
- PMID: 21983565
- PMCID: PMC3208083
- DOI: 10.1038/ncb2354
Microtubules induce self-organization of polarized PAR domains in Caenorhabditis elegans zygotes
Abstract
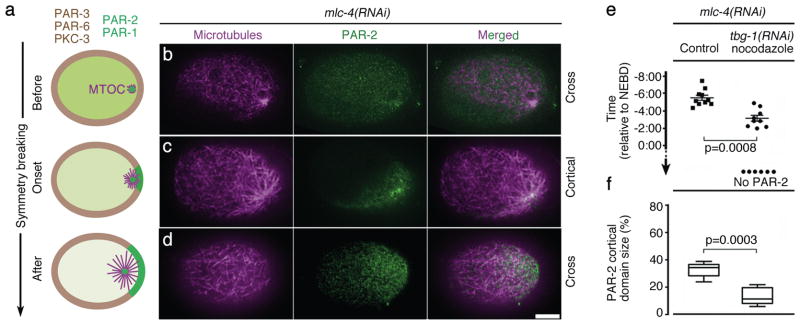
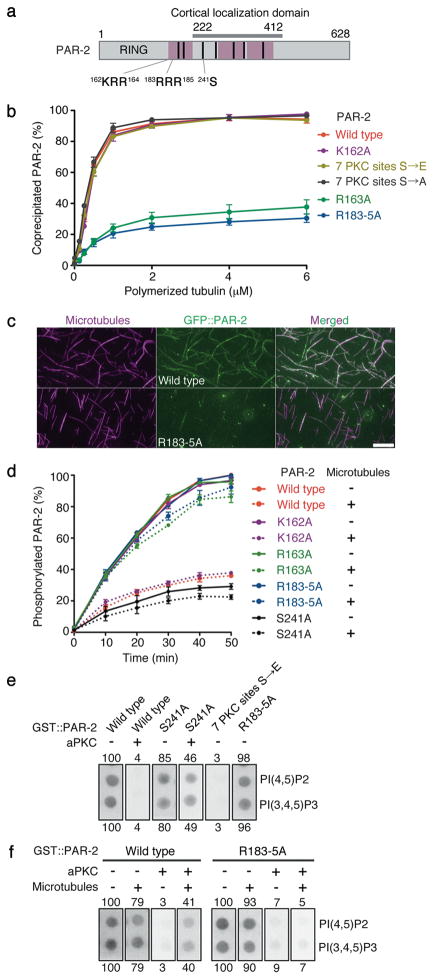
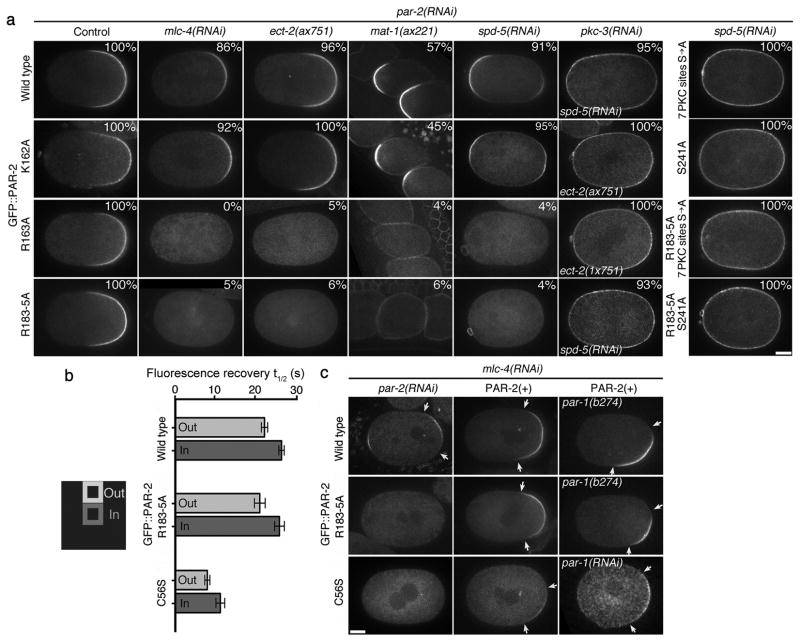
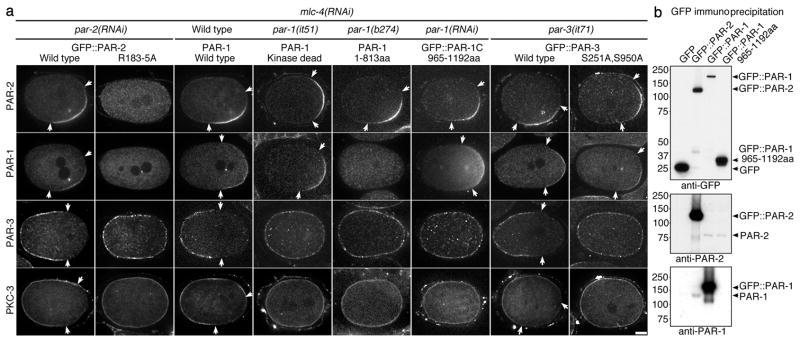
A hallmark of polarized cells is the segregation of the PAR polarity regulators into asymmetric domains at the cell cortex. Antagonistic interactions involving two conserved kinases, atypical protein kinase C (aPKC) and PAR-1, have been implicated in polarity maintenance, but the mechanisms that initiate the formation of asymmetric PAR domains are not understood. Here, we describe one pathway used by the sperm-donated centrosome to polarize the PAR proteins in Caenorhabditis elegans zygotes. Before polarization, cortical aPKC excludes PAR-1 kinase and its binding partner PAR-2 by phosphorylation. During symmetry breaking, microtubules nucleated by the centrosome locally protect PAR-2 from phosphorylation by aPKC, allowing PAR-2 and PAR-1 to access the cortex nearest the centrosome. Cortical PAR-1 phosphorylates PAR-3, causing the PAR-3-aPKC complex to leave the cortex. Our findings illustrate how microtubules, independently of actin dynamics, stimulate the self-organization of PAR proteins by providing local protection against a global barrier imposed by aPKC.
Figures
Comment in
-
Going with the flow: an elegant model for symmetry breaking.Dev Cell. 2011 Dec 13;21(6):981-2. doi: 10.1016/j.devcel.2011.11.015. Dev Cell. 2011. PMID: 22172666
References
-
- St Johnston D, Ahringer J. Cell polarity in eggs and epithelia: parallels and diversity. Cell. 2010;141:757–774. - PubMed
-
- Munro E, Nance J, Priess JR. Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo. Dev Cell. 2004;7:413–424. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
