

Epigenetic suppression of GAD65 expression mediates persistent pain
- PMID: 21983856
- PMCID: PMC3210928
- DOI: 10.1038/nm.2442
Epigenetic suppression of GAD65 expression mediates persistent pain
Abstract
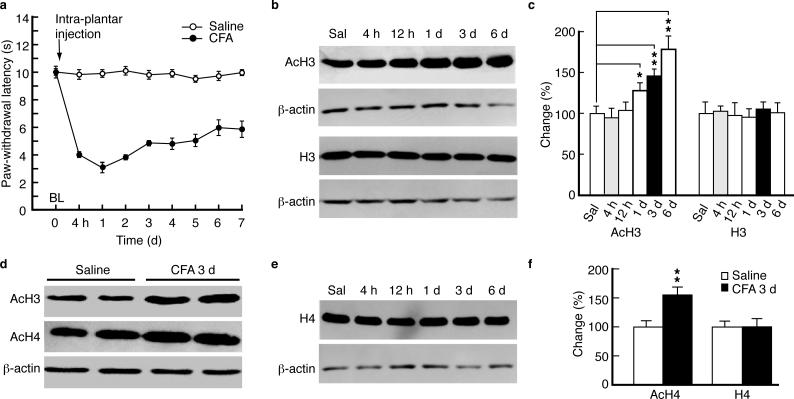
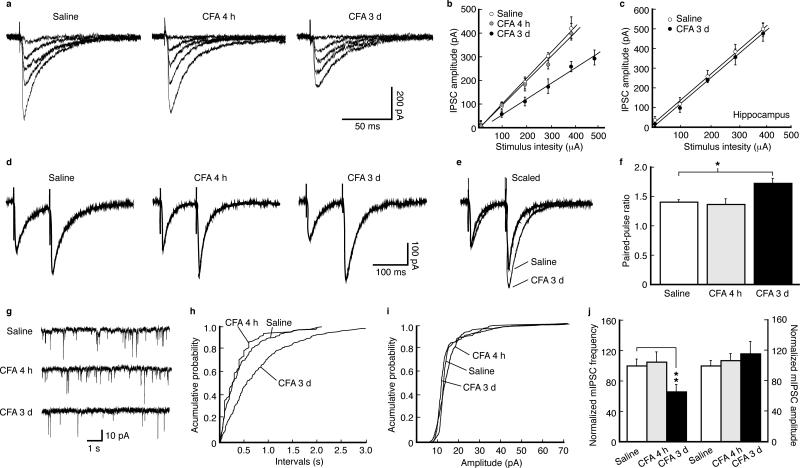
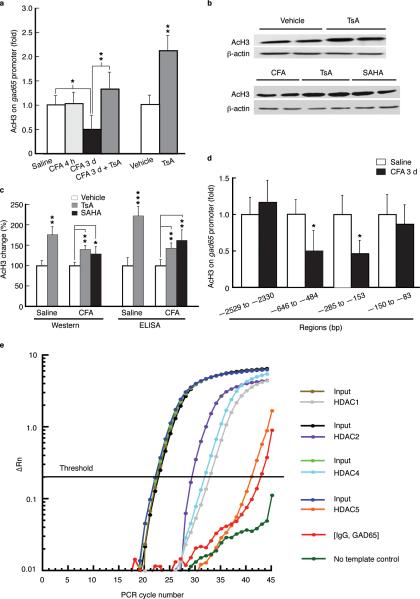
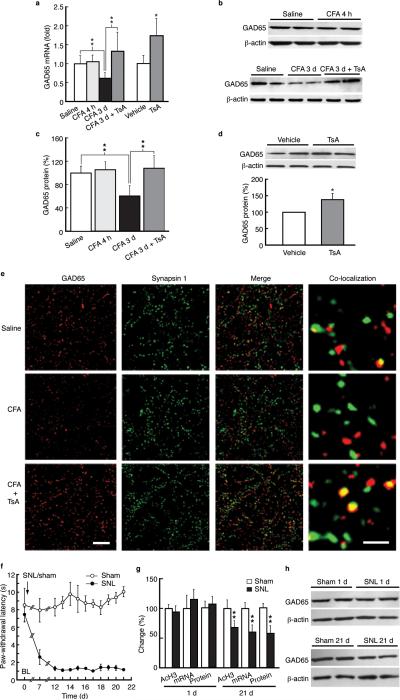
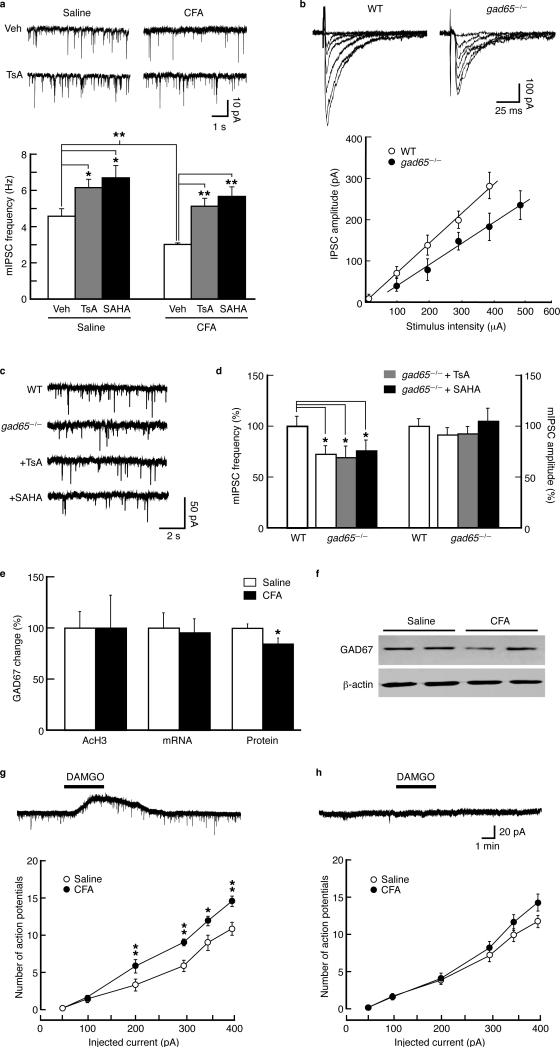
Chronic pain is a common neurological disease involving lasting, multifaceted maladaptations ranging from gene modulation to synaptic dysfunction and emotional disorders. Sustained pathological stimuli in many diseases alter the output activities of certain genes through epigenetic modifications, but it is unclear how epigenetic mechanisms operate in the development of chronic pain. We show here that in the rat brainstem nucleus raphe magnus, which is important for central mechanisms of chronic pain, persistent inflammatory and neuropathic pain epigenetically suppresses Gad2 (encoding glutamic acid decarboxylase 65 (GAD65)) transcription through histone deacetylase (HDAC)-mediated histone hypoacetylation, resulting in impaired γ-aminobutyric acid (GABA) synaptic inhibition. Gad2 knockout mice showed sensitized pain behavior and impaired GABA synaptic function in their brainstem neurons. In wild-type but not Gad2 knockout mice, HDAC inhibitors strongly increased GAD65 activity, restored GABA synaptic function and relieved sensitized pain behavior. These findings suggest GAD65 and HDACs as potential therapeutic targets in an epigenetic approach to the treatment of chronic pain.
Figures

Comment in
-
Analgesia: Unravelling epigenetic mechanisms of chronic pain.Nat Rev Drug Discov. 2011 Nov 18;10(12):900. doi: 10.1038/nrd3606. Nat Rev Drug Discov. 2011. PMID: 22094871 No abstract available.
Similar articles
-
Brainstem brain-derived neurotrophic factor signaling is required for histone deacetylase inhibitor-induced pain relief.Mol Pharmacol. 2015 Jun;87(6):1035-41. doi: 10.1124/mol.115.098186. Epub 2015 Apr 7. Mol Pharmacol. 2015. PMID: 25852071
-
Histone deacetylase inhibitor-induced emergence of synaptic δ-opioid receptors and behavioral antinociception in persistent neuropathic pain.Neuroscience. 2016 Dec 17;339:54-63. doi: 10.1016/j.neuroscience.2016.09.015. Epub 2016 Sep 17. Neuroscience. 2016. PMID: 27646288
-
Transcriptional control of Gad2.Transcription. 2012 Mar-Apr;3(2):68-72. doi: 10.4161/trns.19511. Epub 2012 Mar 1. Transcription. 2012. PMID: 22414751 Free PMC article. Review.
-
Rats deficient in the GAD65 isoform exhibit epilepsy and premature lethality.FASEB J. 2021 Feb;35(2):e21224. doi: 10.1096/fj.202001935R. Epub 2020 Nov 25. FASEB J. 2021. PMID: 33236473
-
[Epigenetic mechanisms and alcohol use disorders: a potential therapeutic target].Biol Aujourdhui. 2017;211(1):83-91. doi: 10.1051/jbio/2017014. Epub 2017 Jul 6. Biol Aujourdhui. 2017. PMID: 28682229 Review. French.
Cited by
-
Improvements in impaired GABA and GAD65/67 production in the spinal dorsal horn contribute to exercise-induced hypoalgesia in a mouse model of neuropathic pain.Mol Pain. 2016 Mar 7;12:1744806916629059. doi: 10.1177/1744806916629059. Print 2016. Mol Pain. 2016. PMID: 27030712 Free PMC article.
-
Normalizing HDAC2 Levels in the Spinal Cord Alleviates Thermal and Mechanical Hyperalgesia After Peripheral Nerve Injury and Promotes GAD65 and KCC2 Expression.Front Neurosci. 2019 Apr 10;13:346. doi: 10.3389/fnins.2019.00346. eCollection 2019. Front Neurosci. 2019. PMID: 31024248 Free PMC article.
-
HDAC inhibitors restore C-fibre sensitivity in experimental neuropathic pain model.Br J Pharmacol. 2013 Nov;170(5):991-8. doi: 10.1111/bph.12366. Br J Pharmacol. 2013. PMID: 24032674 Free PMC article.
-
Evaluation of photobiomodulation therapy (117 and 90s) on pain, regeneration, and epigenetic factors (HDAC 2, DNMT3a) expression following spinal cord injury in a rat model.Photochem Photobiol Sci. 2023 Nov;22(11):2527-2540. doi: 10.1007/s43630-023-00467-5. Epub 2023 Oct 3. Photochem Photobiol Sci. 2023. PMID: 37787959
-
Compensatory Activation of Cannabinoid CB2 Receptor Inhibition of GABA Release in the Rostral Ventromedial Medulla in Inflammatory Pain.J Neurosci. 2017 Jan 18;37(3):626-636. doi: 10.1523/JNEUROSCI.1310-16.2016. J Neurosci. 2017. PMID: 28100744 Free PMC article.
References
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- MacDonald JL, Roskams AJ. Epigenetic regulation of nervous system development by DNA methylation and histone deacetylation. Prog. Neurobiol. 2009;88:170–183. - PubMed
-
- Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. - PubMed
-
- Strahl BD, Allis CD. The language of covalent histone modifications. Nature. 2000;403:41–45. - PubMed
-
- Urdinguio RG, Sanchez-Mut JV, Esteller M. Epigenetic mechanisms in neurological diseases: genes, syndromes, and therapies. Lancet Neurol. 2009;8:1056–1072. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
