

Molecular architecture of a multifunctional MCM complex
- PMID: 21984415
- PMCID: PMC3273815
- DOI: 10.1093/nar/gkr831
Molecular architecture of a multifunctional MCM complex
Abstract
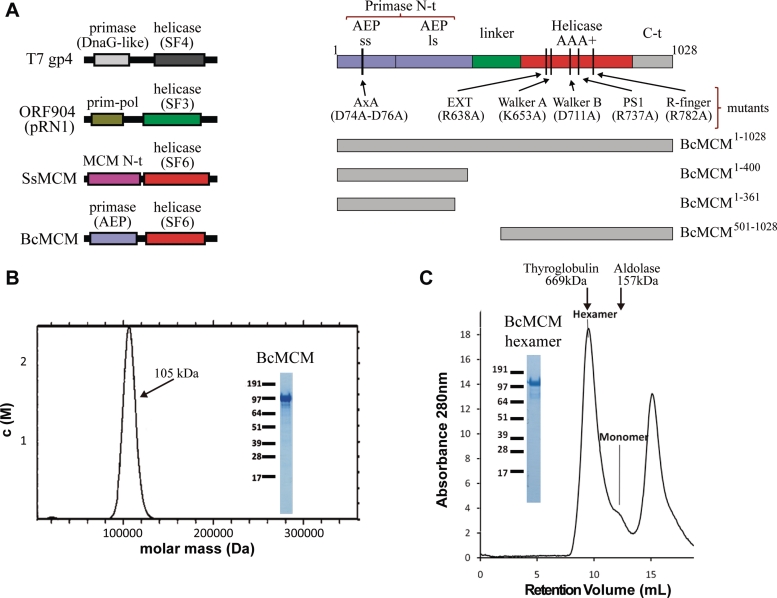
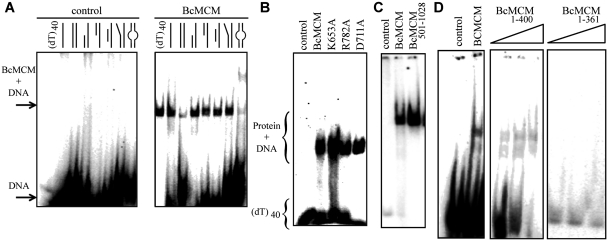
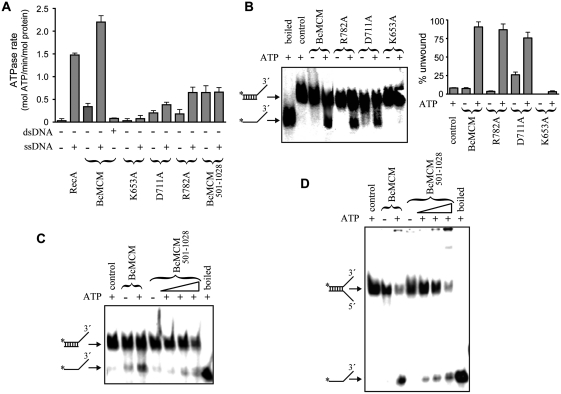
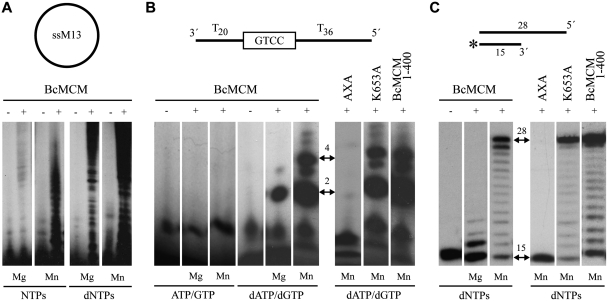
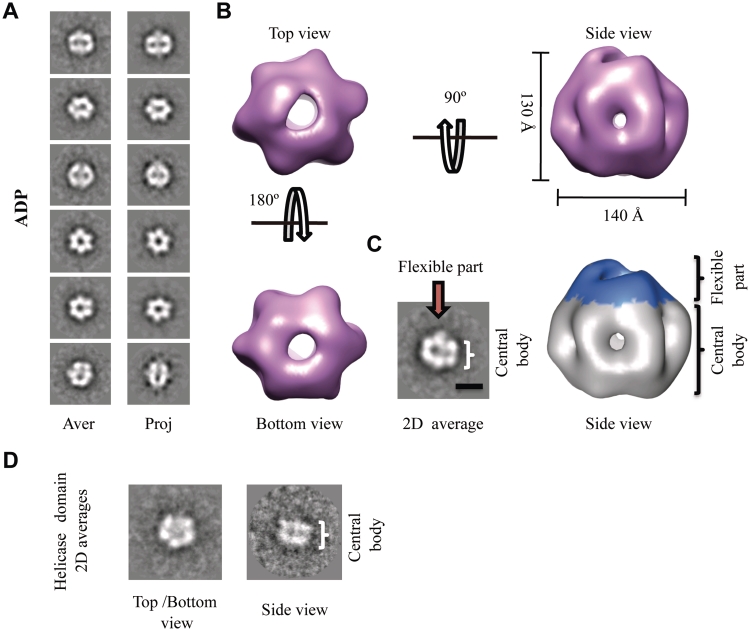
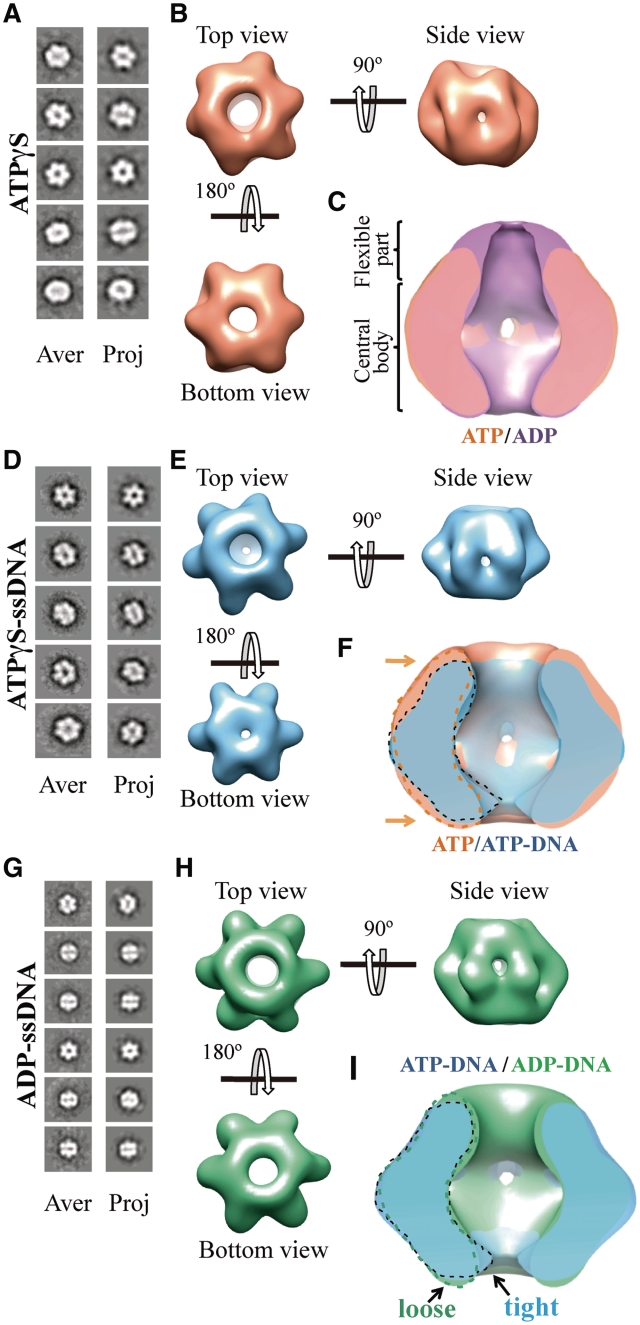
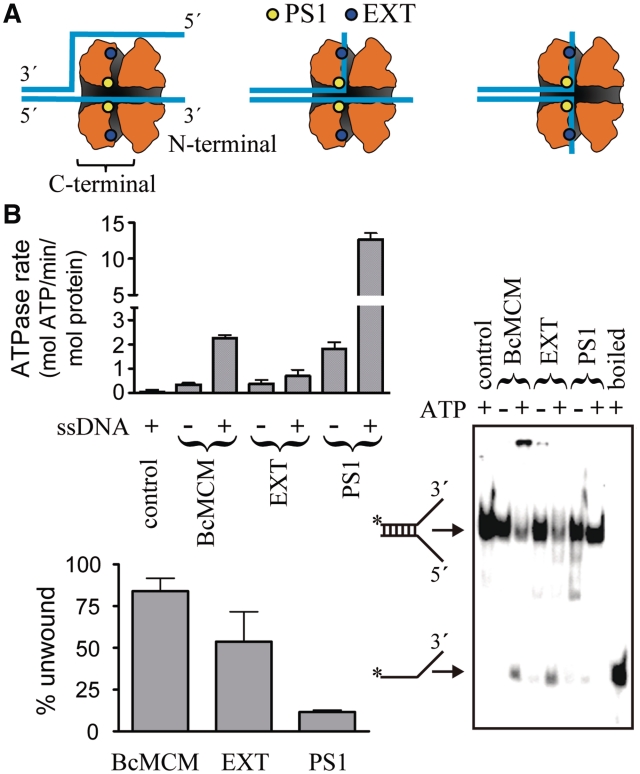
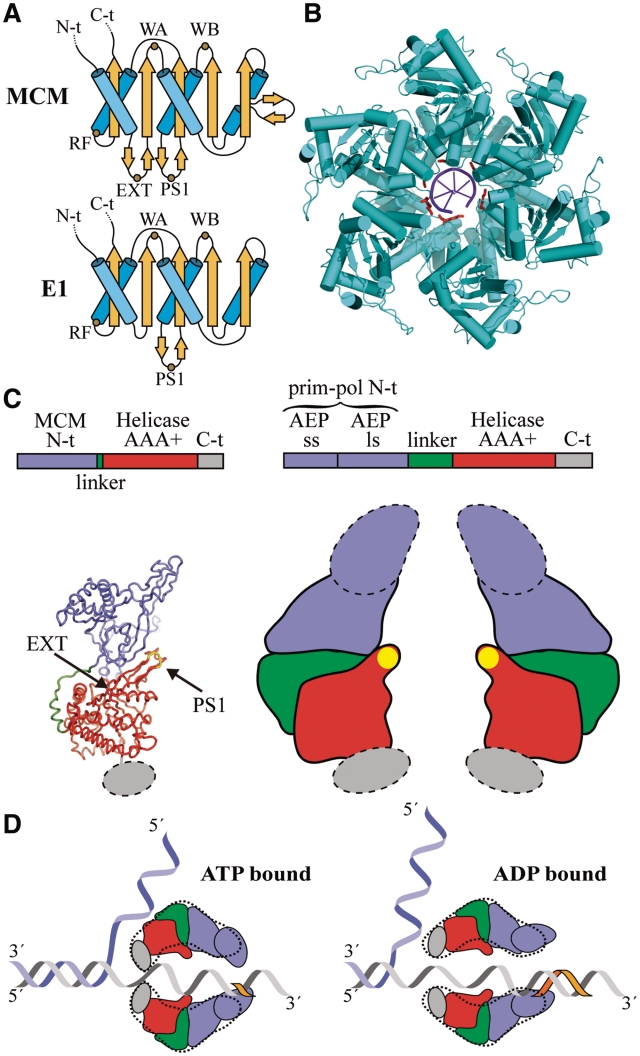
DNA replication is strictly regulated through a sequence of steps that involve many macromolecular protein complexes. One of them is the replicative helicase, which is required for initiation and elongation phases. A MCM helicase found as a prophage in the genome of Bacillus cereus is fused with a primase domain constituting an integrative arrangement of two essential activities for replication. We have isolated this helicase-primase complex (BcMCM) showing that it can bind DNA and displays not only helicase and primase but also DNA polymerase activity. Using single-particle electron microscopy and 3D reconstruction, we obtained structures of BcMCM using ATPγS or ADP in the absence and presence of DNA. The complex depicts the typical hexameric ring shape. The dissection of the unwinding mechanism using site-directed mutagenesis in the Walker A, Walker B, arginine finger and the helicase channels, suggests that the BcMCM complex unwinds DNA following the extrusion model similarly to the E1 helicase from papillomavirus.
Figures
References
-
- Shechter DF, Ying CY, Gautier J. The intrinsic DNA helicase activity of Methanobacterium thermoautotrophicum delta H minichromosome maintenance protein. J. Biol. Chem. 2000;275:15049–15059. - PubMed
-
- Niedenzu T, Roleke D, Bains G, Scherzinger E, Saenger W. Crystal structure of the hexameric replicative helicase RepA of plasmid RSF1010. J. Mol. Biol. 2001;306:479–487. - PubMed
-
- Bochman ML, Schwacha A. The Mcm2-7 complex has in vitro helicase activity. Mol. Cell. 2008;31:287–293. - PubMed
-
- Gomez-Llorente Y, Fletcher RJ, Chen XS, Carazo JM, San Martin C. Polymorphism and double hexamer structure in the archaeal minichromosome maintenance (MCM) helicase from Methanobacterium thermoautotrophicum. J. Biol. Chem. 2005;280:40909–40915. - PubMed
