

Unique properties of the ATP-sensitive K⁺ channel in the mouse ventricular cardiac conduction system
- PMID: 21984445
- PMCID: PMC3247655
- DOI: 10.1161/CIRCEP.111.964643
Unique properties of the ATP-sensitive K⁺ channel in the mouse ventricular cardiac conduction system
Abstract
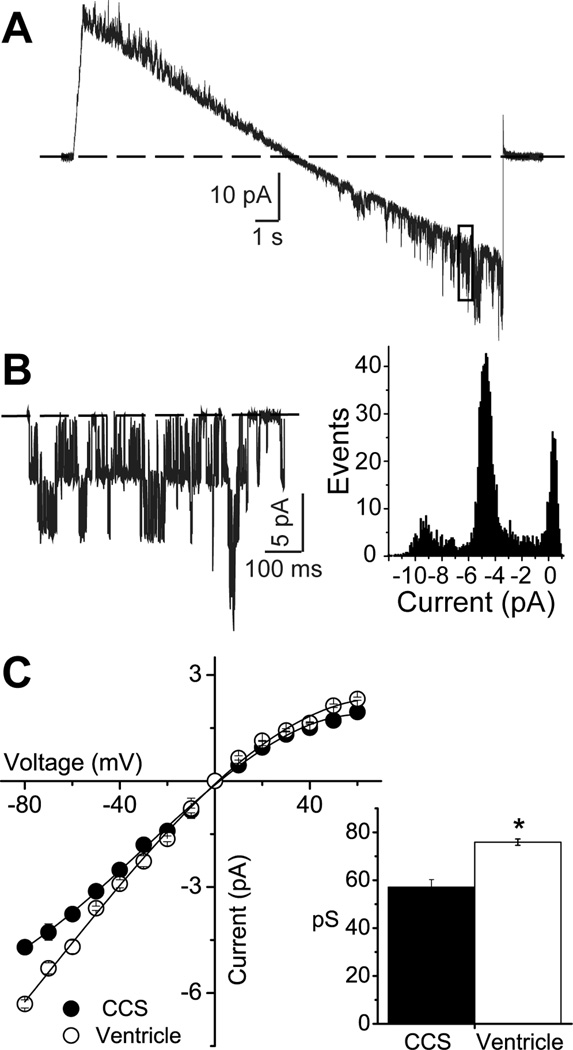
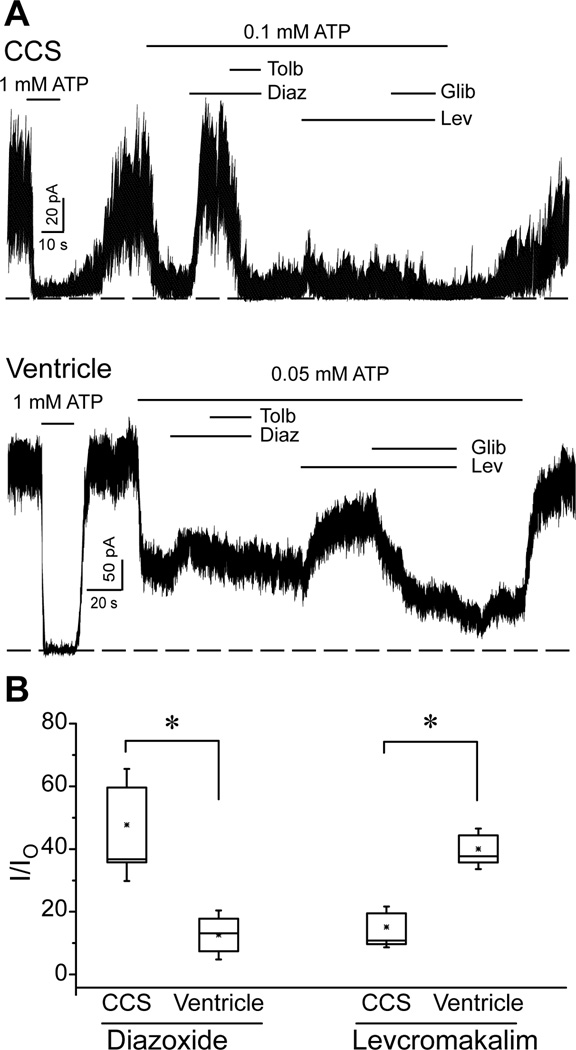
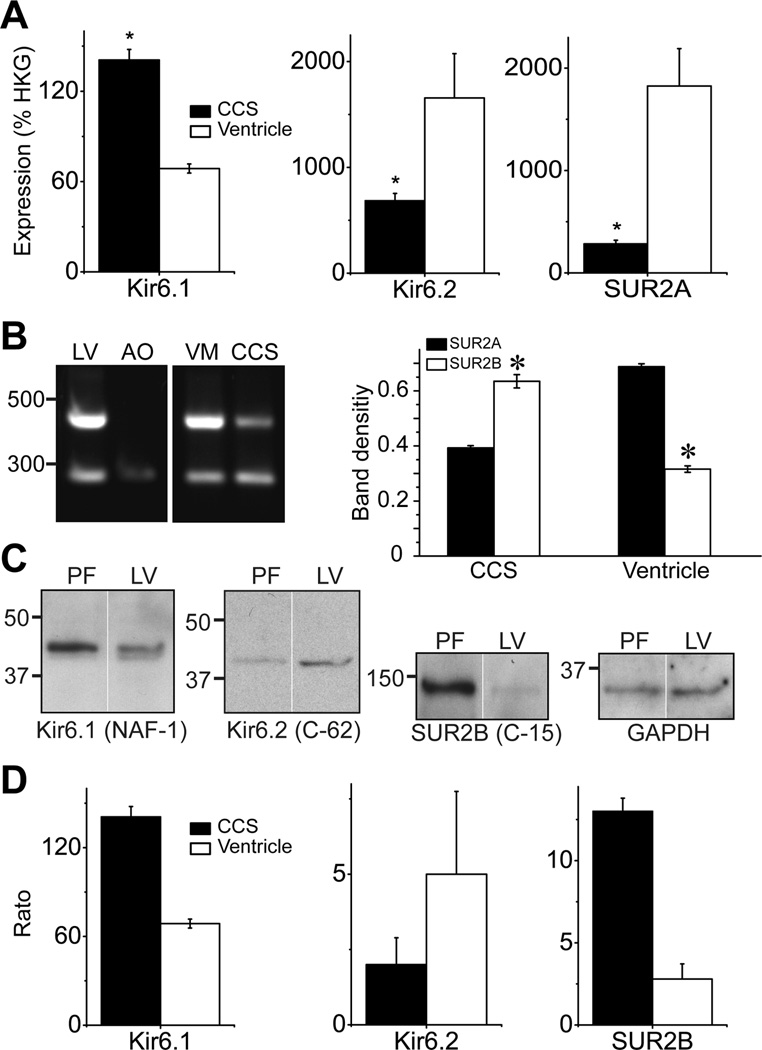
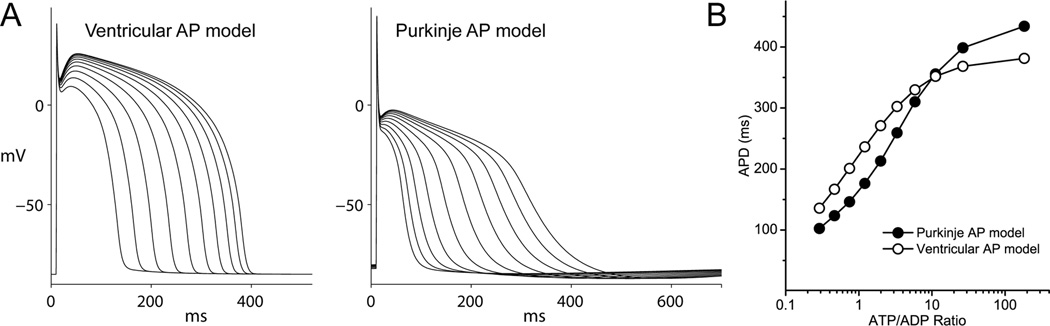
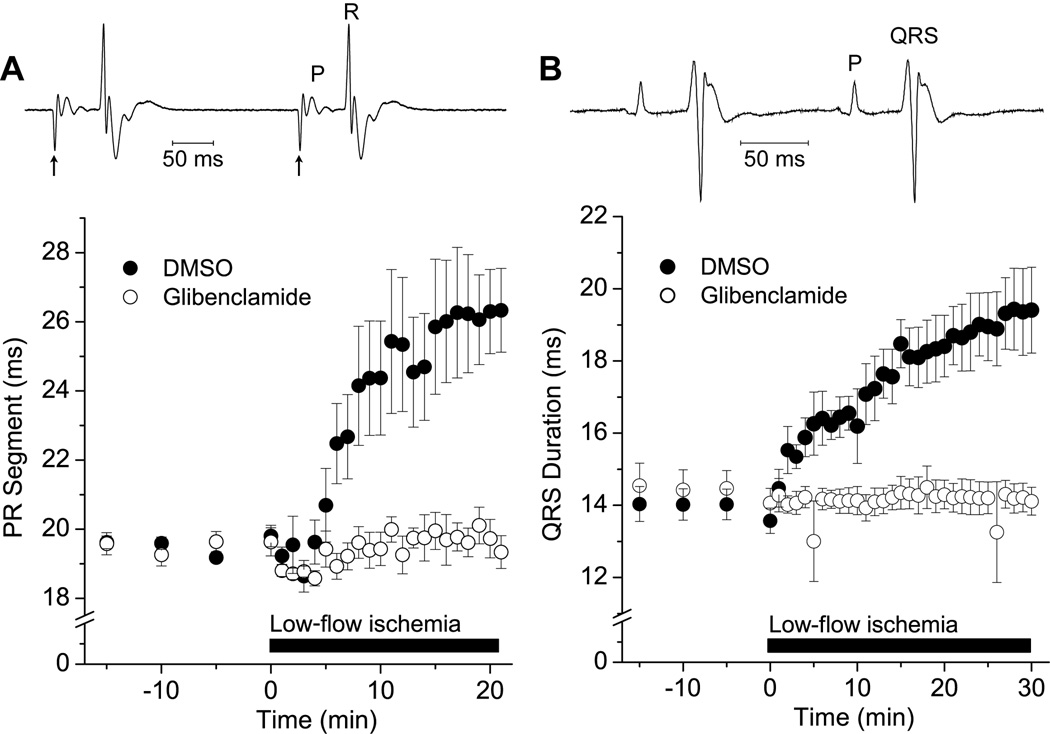
Background- The specialized cardiac conduction system (CCS) expresses a unique complement of ion channels that confer a specific electrophysiological profile. ATP-sensitive potassium (K(ATP)) channels in these myocytes have not been systemically investigated. Methods and Results- We recorded K(ATP) channels in isolated CCS myocytes using Cntn2-EGFP reporter mice. The CCS K(ATP) channels were less sensitive to inhibitory cytosolic ATP compared with ventricular channels and more strongly activated by MgADP. They also had a smaller slope conductance. The 2 types of channels had similar intraburst open and closed times, but the CCS K(ATP) channel had a prolonged interburst closed time. CCS K(ATP) channels were strongly activated by diazoxide and less by levcromakalim, whereas the ventricular K(ATP) channel had a reverse pharmacological profile. CCS myocytes express elevated levels of Kir6.1 but reduced Kir6.2 and SUR2A mRNA compared with ventricular myocytes (SUR1 expression was negligible). SUR2B mRNA expression was higher in CCS myocytes relative to SUR2A. Canine Purkinje fibers expressed higher levels of Kir6.1 and SUR2B protein relative to the ventricle. Numeric simulation predicts a high sensitivity of the Purkinje action potential to changes in ATP:ADP ratio. Cardiac conduction time was prolonged by low-flow ischemia in isolated, perfused mouse hearts, which was prevented by glibenclamide. Conclusions- These data imply a differential electrophysiological response (and possible contribution to arrhythmias) of the ventricular CCS to K(ATP) channel opening during periods of ischemia.
Conflict of interest statement
Figures


Comment in
-
"Cardiac KATP": a family of ion channels.Circ Arrhythm Electrophysiol. 2011 Dec;4(6):796-8. doi: 10.1161/CIRCEP.111.968081. Circ Arrhythm Electrophysiol. 2011. PMID: 22203659 Free PMC article. No abstract available.
References
-
- Noble D. The initiation of the heart beat. Oxford: Oxford University Press; 1975.
-
- Janse MJ, Kleber AG. Electrophysiological changes and ventricular arrhythmias in the early phase of regional myocardial ischemia. Circulation Research. 1981;49:1069–1081. - PubMed
-
- Noma A. ATP-regulated K+ channels in cardiac muscle. Nature. 1983;305:147–148. - PubMed
-
- Ito H, Nakajima T, Takikawa R, Hamada E, Iguchi M, Sugimoto T, Kurachi Y. Coenzyme Q10 attenuates cyanide-activation of the ATP-sensitive K+ channel current in single cardiac myocytes of the guinea-pig. Naunyn Schmiedebergs Arch Pharmacol. 1991;344:133–136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL081336/HL/NHLBI NIH HHS/United States
- HL076230/HL/NHLBI NIH HHS/United States
- R01 HL076230/HL/NHLBI NIH HHS/United States
- HL105046/HL/NHLBI NIH HHS/United States
- R01 HL076751/HL/NHLBI NIH HHS/United States
- HL82727/HL/NHLBI NIH HHS/United States
- R01 HL105983/HL/NHLBI NIH HHS/United States
- HL076751/HL/NHLBI NIH HHS/United States
- R01 HL082727/HL/NHLBI NIH HHS/United States
- F32 HL105046/HL/NHLBI NIH HHS/United States
- HL093563/HL/NHLBI NIH HHS/United States
- R21 HL093563/HL/NHLBI NIH HHS/United States
- HL105983/HL/NHLBI NIH HHS/United States
- HL085820/HL/NHLBI NIH HHS/United States
- R01 HL085820/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
