

The coronary circulation in exercise training
- PMID: 21984538
- PMCID: PMC3334245
- DOI: 10.1152/ajpheart.00574.2011
The coronary circulation in exercise training
Abstract
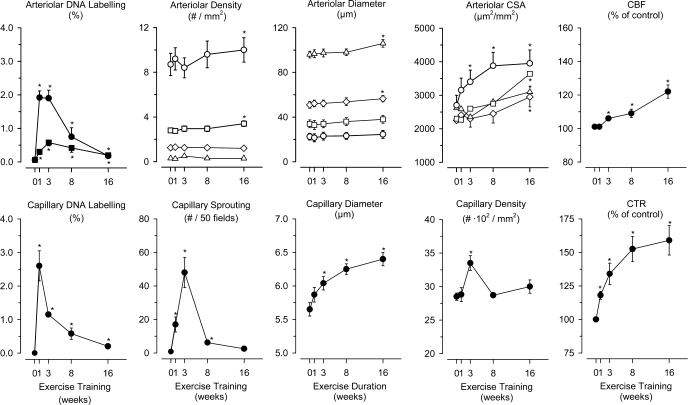
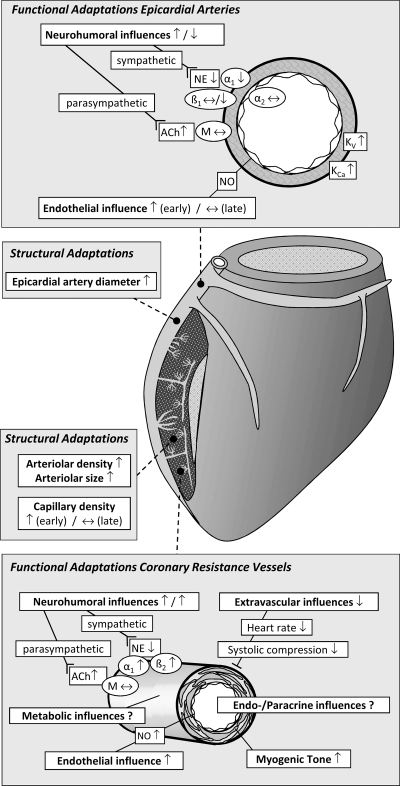
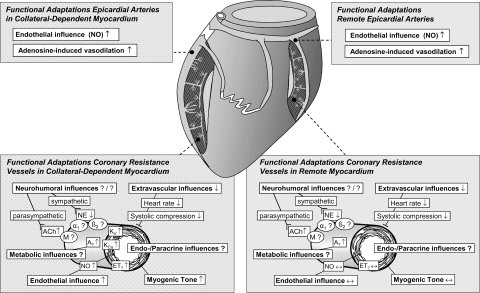
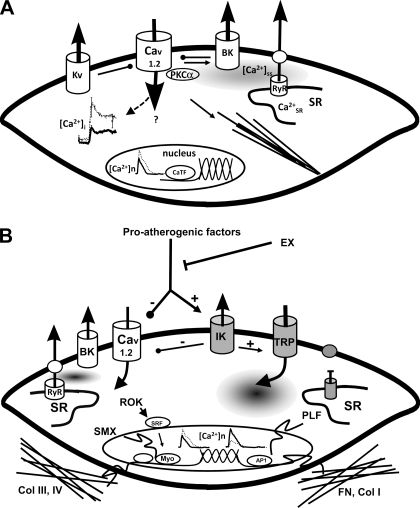
Exercise training (EX) induces increases in coronary transport capacity through adaptations in the coronary microcirculation including increased arteriolar diameters and/or densities and changes in the vasomotor reactivity of coronary resistance arteries. In large animals, EX increases capillary exchange capacity through angiogenesis of new capillaries at a rate matched to EX-induced cardiac hypertrophy so that capillary density remains normal. However, after EX coronary capillary exchange area is greater (i.e., capillary permeability surface area product is greater) at any given blood flow because of altered coronary vascular resistance and matching of exchange surface area and blood flow distribution. The improved coronary capillary blood flow distribution appears to be the result of structural changes in the coronary tree and alterations in vasoreactivity of coronary resistance arteries. EX also alters vasomotor reactivity of conduit coronary arteries in that after EX, α-adrenergic receptor responsiveness is blunted. Of interest, α- and β-adrenergic tone appears to be maintained in the coronary microcirculation in the presence of lower circulating catecholamine levels because of increased receptor responsiveness to adrenergic stimulation. EX also alters other vasomotor control processes of coronary resistance vessels. For example, coronary arterioles exhibit increased myogenic tone after EX, likely because of a calcium-dependent PKC signaling-mediated alteration in voltage-gated calcium channel activity in response to stretch. Conversely, EX augments endothelium-dependent vasodilation throughout the coronary arteriolar network and in the conduit arteries in coronary artery disease (CAD). The enhanced endothelium-dependent dilation appears to result from increased nitric oxide bioavailability because of changes in nitric oxide synthase expression/activity and decreased oxidant stress. EX also decreases extravascular compressive forces in the myocardium at rest and at comparable levels of exercise, mainly because of decreases in heart rate and duration of systole. EX does not stimulate growth of coronary collateral vessels in the normal heart. However, if exercise produces ischemia, which would be absent or minimal under resting conditions, there is evidence that collateral growth can be enhanced. While there is evidence that EX can decrease the progression of atherosclerotic lesions or even induce the regression of atherosclerotic lesions in humans, the evidence of this is not strong due to the fact that most prospective trials conducted to date have included other lifestyle changes and treatment strategies by necessity. The literature from large animal models of CAD also presents a cloudy picture concerning whether EX can induce the regression of or slow the progression of atherosclerotic lesions. Thus, while evidence from research using humans with CAD and animal models of CAD indicates that EX increases endothelium-dependent dilation throughout the coronary vascular tree, evidence that EX reverses or slows the progression of lesion development in CAD is not conclusive at this time. This suggests that the beneficial effects of EX in CAD may not be the result of direct effects on the coronary artery wall. If this suggestion is true, it is important to determine the mechanisms involved in these beneficial effects.
Figures
References
-
- Anversa P, Levicky V, Beghi C, McDonald SL, Kikkawa Y. Morphometry of exercise-induced right ventricular hypertrophy in the rat. Circ Res 52: 57–64, 1983 - PubMed
-
- Bache RJ. Effects of hypertrophy on the cornary circulation. Prog Cardiovasc Dis 30: 403–440, 1988 - PubMed
-
- Bache RJ, Dai XZ, Schwartz JS, Homans DC. Role of adenosine in coronary vasodilation during exercise. Circ Res 62: 846–853, 1988 - PubMed
-
- Barbier J, Reland S, Ville N, Rannou-Bekono F, Wong S, Carre F. The effects of exercise training on myocardial adrenergic and muscarinic receptors. Clin Auton Res 16: 61–65, 2006 - PubMed
-
- Barnard RJ, Duncan HW, Baldwin KM, Grimditch G, Buckberg DD. Effects of intensive exercise training on myocardial performance and coronary blood flow. J Appl Physiol 49: 444–449, 1980 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous