

BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development
- PMID: 21984813
- PMCID: PMC3196857
- DOI: 10.1242/jcs.083659
BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development
Abstract
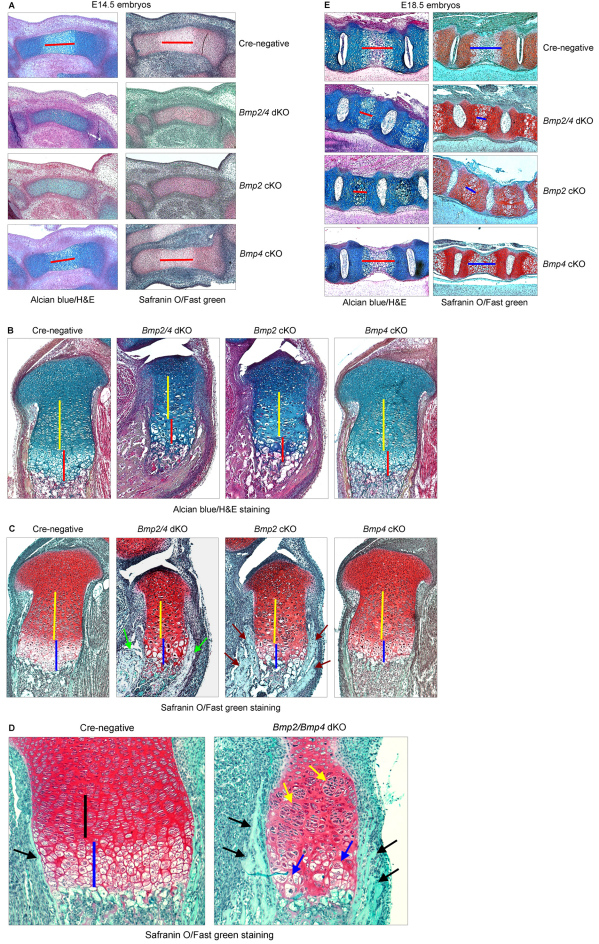
The BMP signaling pathway has a crucial role in chondrocyte proliferation and maturation during endochondral bone development. To investigate the specific function of the Bmp2 and Bmp4 genes in growth plate chondrocytes during cartilage development, we generated chondrocyte-specific Bmp2 and Bmp4 conditional knockout (cKO) mice and Bmp2,Bmp4 double knockout (dKO) mice. We found that deletion of Bmp2 and Bmp4 genes or the Bmp2 gene alone results in a severe chondrodysplasia phenotype, whereas deletion of the Bmp4 gene alone produces a minor cartilage phenotype. Both dKO and Bmp2 cKO mice exhibit severe disorganization of chondrocytes within the growth plate region and display profound defects in chondrocyte proliferation, differentiation and apoptosis. To understand the mechanism by which BMP2 regulates these processes, we explored the specific relationship between BMP2 and Runx2, a key regulator of chondrocyte differentiation. We found that BMP2 induces Runx2 expression at both the transcriptional and post-transcriptional levels. BMP2 enhances Runx2 protein levels through inhibition of CDK4 and subsequent prevention of Runx2 ubiquitylation and proteasomal degradation. Our studies provide novel insights into the genetic control and molecular mechanism of BMP signaling during cartilage development.
Figures





References
-
- Alden T. D., Pittman D. D., Hankins G. R., Beres E. J., Engh J. A., Das S., Hudson S. B., Kerns K. M., Kallmes D. F., Helm G. A. (1999). In vivo endochondral bone formation using a bone morphogenetic protein 2 adenoviral vector. Human Gene. Ther. 10, 2245-2253 - PubMed
-
- Chen D., Harris M. A., Rossini G., Dunstan C. R., Dallas S. L., Feng J. Q., Mundy G. R., Harris S. E. (1997). Bone morphogenetic protein 2 (BMP-2) enhances BMP-3, BMP-4 and bone cell differentiation marker gene expression during the induction of mineralized bone matrix formation in cultures of fetal rat calvarial osteoblasts. Calcif. Tissue Int. 60, 283-290 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
