

ATP synthase complex of Plasmodium falciparum: dimeric assembly in mitochondrial membranes and resistance to genetic disruption
- PMID: 21984828
- PMCID: PMC3308843
- DOI: 10.1074/jbc.M111.290973
ATP synthase complex of Plasmodium falciparum: dimeric assembly in mitochondrial membranes and resistance to genetic disruption
Abstract
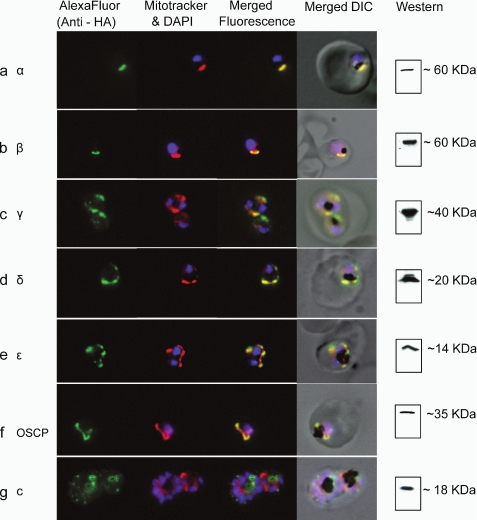
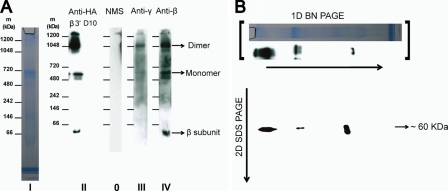
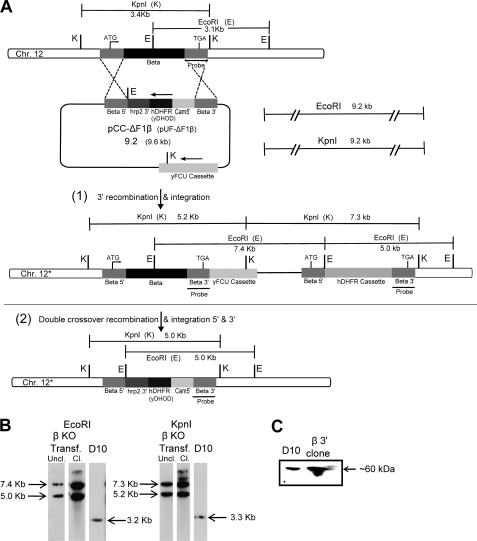
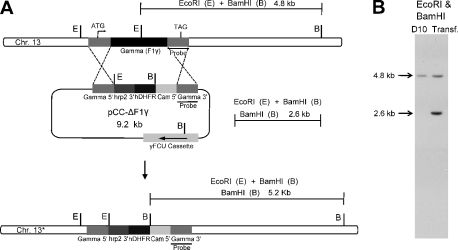
The rotary nanomotor ATP synthase is a central player in the bioenergetics of most organisms. Yet the role of ATP synthase in malaria parasites has remained unclear, as blood stages of Plasmodium falciparum appear to derive ATP largely through glycolysis. Also, genes for essential subunits of the F(O) sector of the complex could not be detected in the parasite genomes. Here, we have used molecular genetic and immunological tools to investigate the localization, complex formation, and functional significance of predicted ATP synthase subunits in P. falciparum. We generated transgenic P. falciparum lines expressing seven epitope-tagged canonical ATP synthase subunits, revealing localization of all but one of the subunits to the mitochondrion. Blue native gel electrophoresis of P. falciparum mitochondrial membranes suggested the molecular mass of the ATP synthase complex to be greater than 1 million daltons. This size is consistent with the complex being assembled as a dimer in a manner similar to the complexes observed in other eukaryotic organisms. This observation also suggests the presence of previously unknown subunits in addition to the canonical subunits in P. falciparum ATP synthase complex. Our attempts to disrupt genes encoding β and γ subunits were unsuccessful, suggesting an essential role played by the ATP synthase complex in blood stages of P. falciparum. These studies suggest that, despite some unconventional features and its minimal contribution to ATP synthesis, P. falciparum ATP synthase is localized to the parasite mitochondrion, assembled as a large dimeric complex, and is likely essential for parasite survival.
Figures
References
-
- Walker J. E., Dickson V. K. (2006) Biochim. Biophys. Acta 1757, 286–296 - PubMed
-
- Devenish R. J., Prescott M., Rodgers A. J. (2008) Int. Rev. Cell Mol. Biol. 267, 1–58 - PubMed
-
- Angevine C. M., Herold K. A., Vincent O. D., Fillingame R. H. (2007) J. Biol. Chem. 282, 9001–9007 - PubMed
-
- Fry M., Beesley J. E. (1991) Parasitology 102, 17–26 - PubMed
-
- Krungkrai J., Burat D., Kudan S., Krungkrai S., Prapunwattana P. (1999) Southeast Asian J. Trop. Med. Public Health 30, 636–642 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
