

Starvation compromises Paneth cells
- PMID: 21986443
- PMCID: PMC3260859
- DOI: 10.1016/j.ajpath.2011.08.030
Starvation compromises Paneth cells
Abstract
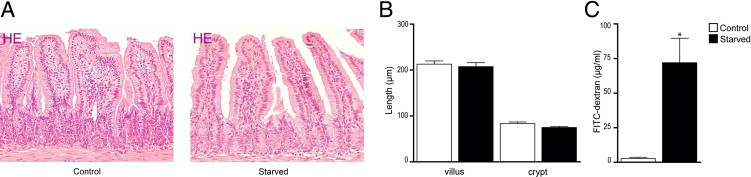
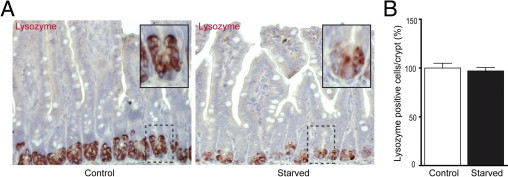
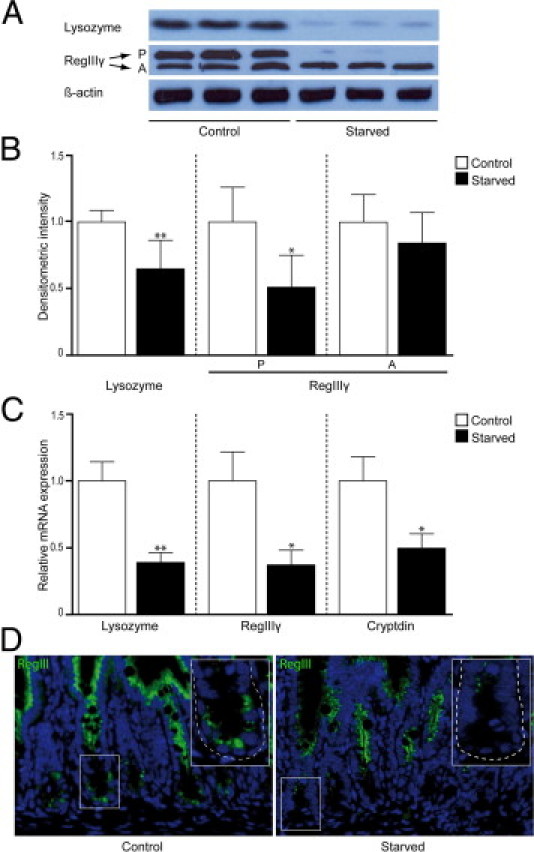
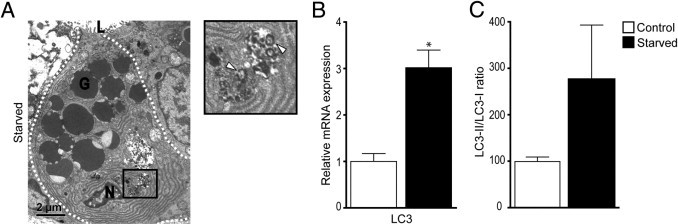
Lack of enteral feeding, with or without parenteral nutritional support, is associated with increased intestinal permeability and translocation of bacteria. Such translocation is thought to be important in the high morbidity and mortality rates of patients who receive nothing by mouth. Recently, Paneth cells, important constituents of innate intestinal immunity, were found to be crucial in host protection against invasion of both commensal and pathogenic bacteria. This study investigates the influence of food deprivation on Paneth cell function in a mouse starvation model. Quantitative PCR showed significant decreases in mRNA expression of typical Paneth cell antimicrobials, lysozyme, cryptdin, and RegIIIγ, in ileal tissue after 48 hours of food deprivation. Protein expression levels of lysozyme and RegIIIγ precursor were also significantly diminished, as shown by Western blot analysis and IHC. Late degenerative autophagolysosomes and aberrant Paneth cell granules in starved mice were evident by electron microscopy, Western blot analysis, and quantitative PCR. Furthermore, increased bacterial translocation to mesenteric lymph nodes coincided with Paneth cell abnormalities. The current study demonstrates the occurrence of Paneth cell abnormalities during enteral starvation. Such changes may contribute to loss of epithelial barrier function, causing the apparent bacterial translocation in enteral starvation.
Copyright © 2011 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
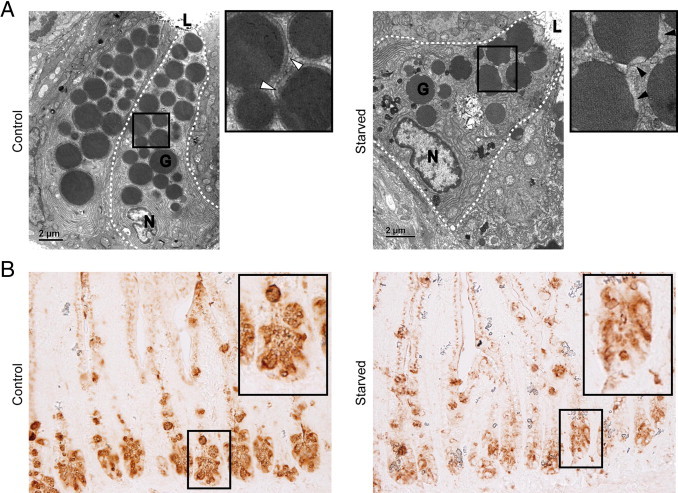
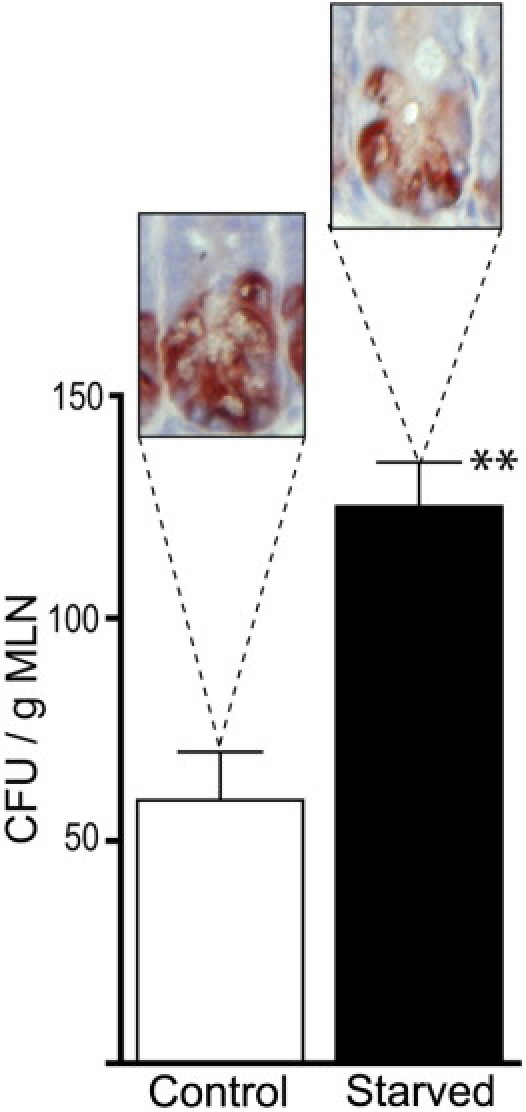
References
-
- Bevins C.L., Salzman N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol. 2011;9:356–368. - PubMed
-
- Ayabe T., Satchell D.P., Wilson C.L., Parks W.C., Selsted M.E., Ouellette A.J. Secretion of microbicidal alpha-defensins by intestinal Paneth cells in response to bacteria. Nat Immunol. 2000;1:113–118. - PubMed
-
- Grootjans J., Hodin C.M., de Haan J.J., Derikx J.P., Rouschop K.M., Verheyen F.K., van Dam R.M., Dejong C.H., Buurman W.A., Lenaerts K. Level of activation of the unfolded protein response correlates with Paneth cell apoptosis in human small intestine exposed to ischemia/reperfusion. Gastroenterology. 2011;140:529–539. - PubMed
-
- MacFie J. Enteral versus parenteral nutrition: the significance of bacterial translocation and gut-barrier function. Nutrition. 2000;16:606–611. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
