

Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis
- PMID: 21989915
- PMCID: PMC3190384
- DOI: 10.1242/dev.067710
Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis
Abstract
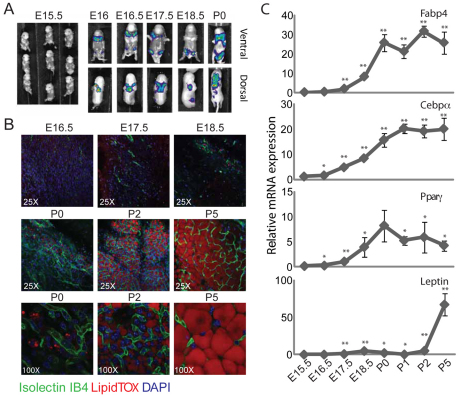
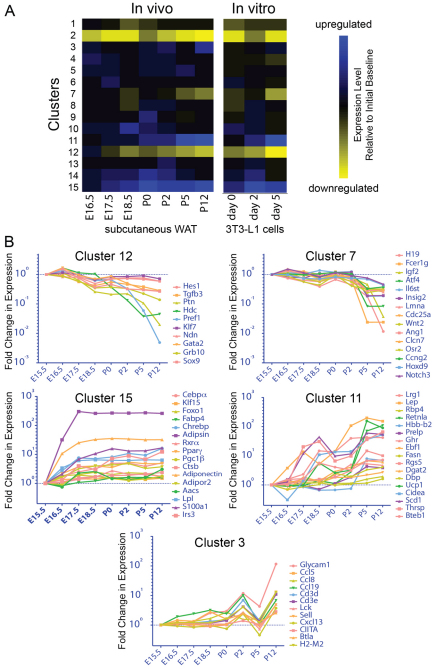
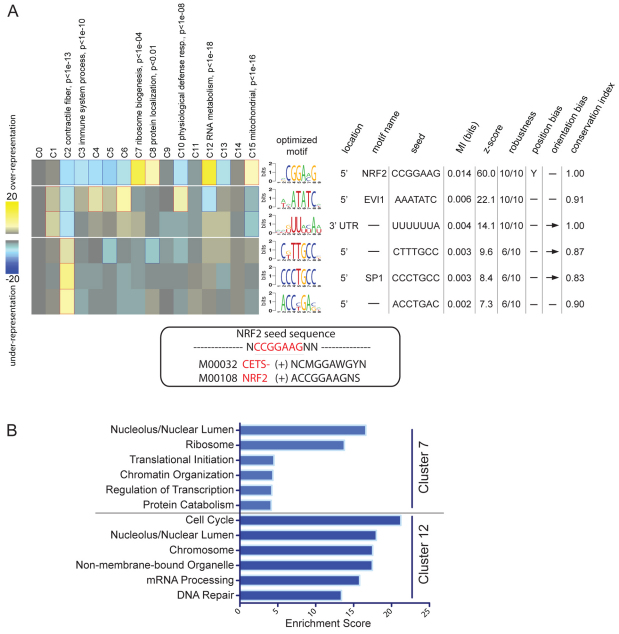
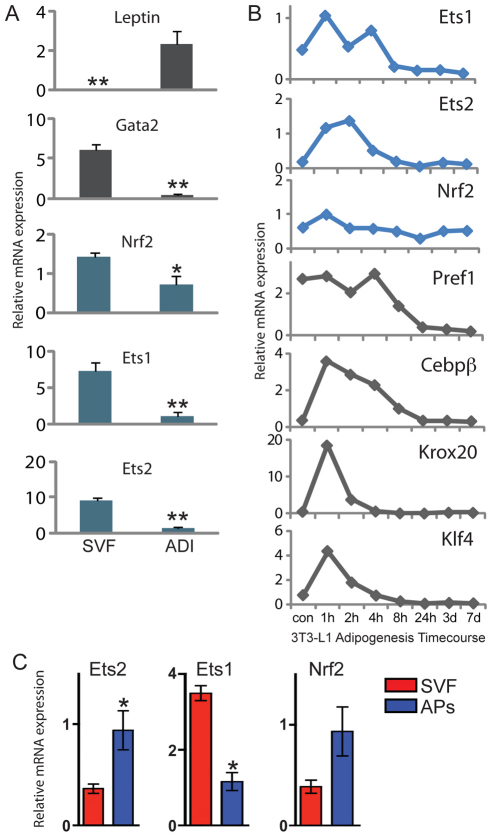
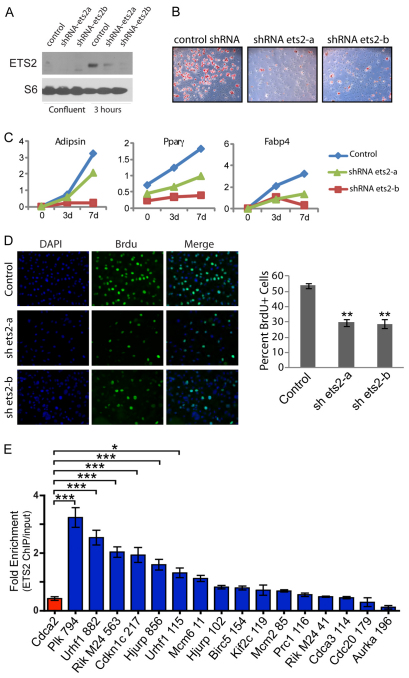
Obesity is characterized by an expansion of white adipose tissue mass that results from an increase in the size and the number of adipocytes. However, the mechanisms responsible for the formation of adipocytes during development and the molecular mechanisms regulating their increase and maintenance in adulthood are poorly understood. Here, we report the use of leptin-luciferase BAC transgenic mice to track white adipose tissue (WAT) development and guide the isolation and molecular characterization of adipocytes during development using DNA microarrays. These data reveal distinct transcriptional programs that are regulated during murine WAT development in vivo. By using a de novo cis-regulatory motif discovery tool (FIRE), we identify two early gene clusters whose promoters show significant enrichment for NRF2/ETS transcription factor binding sites. We further demonstrate that Ets transcription factors, but not Nrf2, are regulated during early adipogenesis and that Ets2 is essential for the normal progression of the adipocyte differentiation program in vitro. These data identify ETS2 as a functionally important transcription factor in adipogenesis and its possible role in regulating adipose tissue mass in adults can now be tested. Our approach also provides the basis for elucidating the function of other gene networks during WAT development in vivo. Finally these data confirm that although gene expression during adipogenesis in vitro recapitulates many of the patterns of gene expression in vivo, there are additional developmental transitions in pre and post-natal adipose tissue that are not evident in cell culture systems.
Figures
References
-
- Barak Y., Nelson M. C., Ong E. S., Jones Y. Z., Ruiz-Lozano P., Chien K. R., Koder A., Evans R. M. (1999). PPAR gamma is required for placental, cardiac, and adipose tissue development. Mol. Cell 4, 585-595 - PubMed
-
- Birkenmeier E. H., Gwynn B., Howard S., Jerry J., Gordon J. I., Landschulz W. H., McKnight S. L. (1989). Tissue-specific expression, developmental regulation, and genetic mapping of the gene encoding CCAAT/enhancer binding protein. Genes Dev. 3, 1146-1156 - PubMed
-
- Bluher M., Michael M. D., Peroni O. D., Ueki K., Carter N., Kahn B. B., Kahn C. R. (2002). Adipose tissue selective insulin receptor knockout protects against obesity and obesity-related glucose intolerance. Dev. Cell 3, 25-38 - PubMed
-
- Chen Z., Torrens J. I., Anand A., Spiegelman B. M., Friedman J. M. (2005). Krox20 stimulates adipogenesis via C/EBPbeta-dependent and -independent mechanisms. Cell Metab. 1, 93-106 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
