

Lipopolysaccharide initiates inflammation in bovine granulosa cells via the TLR4 pathway and perturbs oocyte meiotic progression in vitro
- PMID: 21990308
- PMCID: PMC3428914
- DOI: 10.1210/en.2011-1124
Lipopolysaccharide initiates inflammation in bovine granulosa cells via the TLR4 pathway and perturbs oocyte meiotic progression in vitro
Abstract
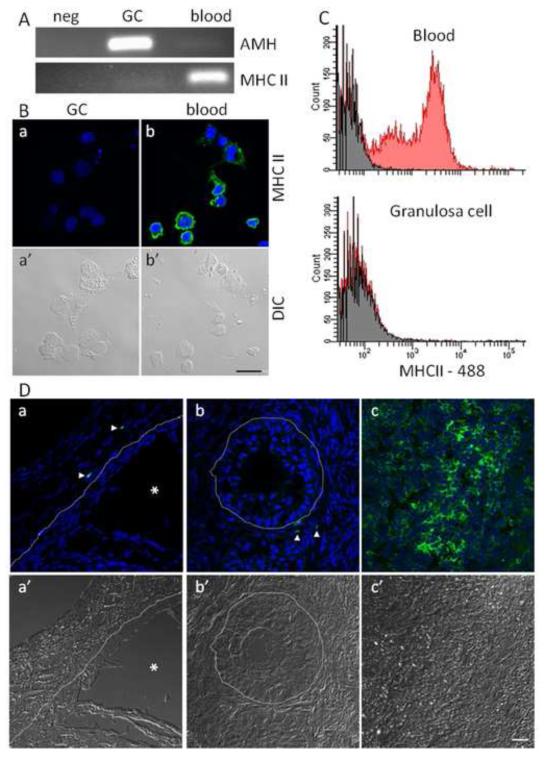
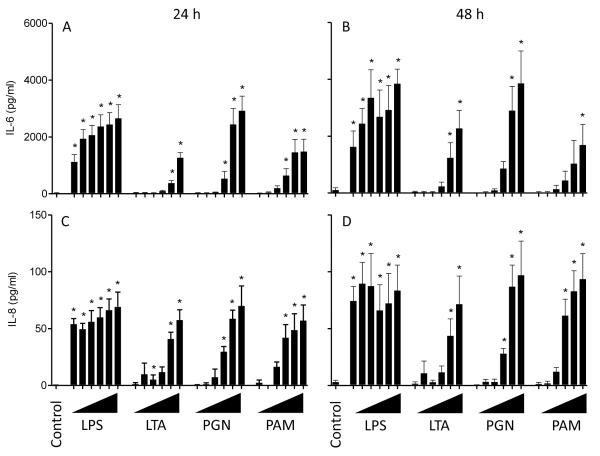
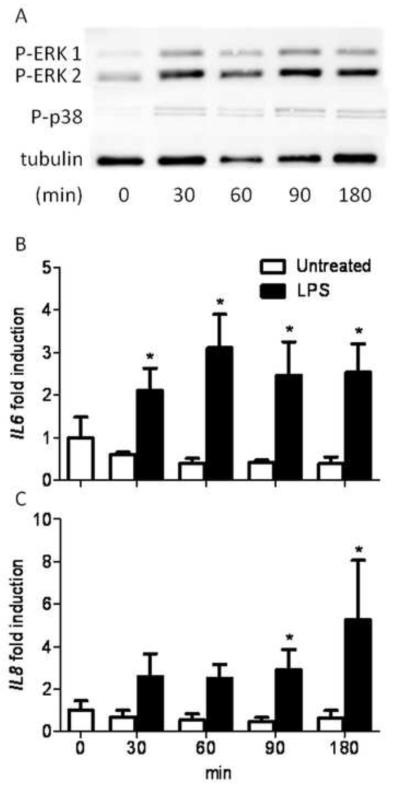
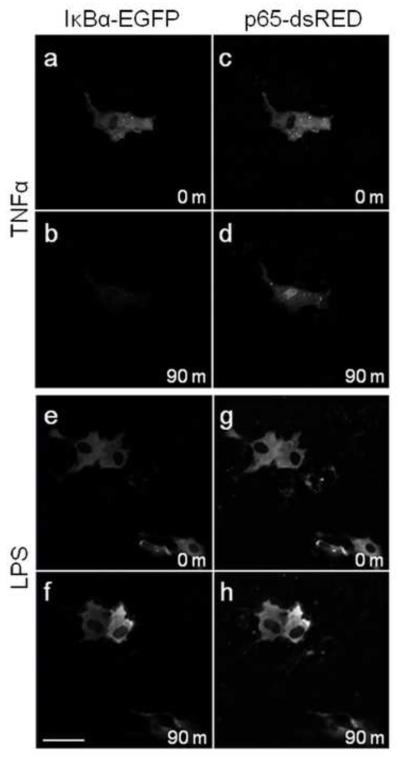
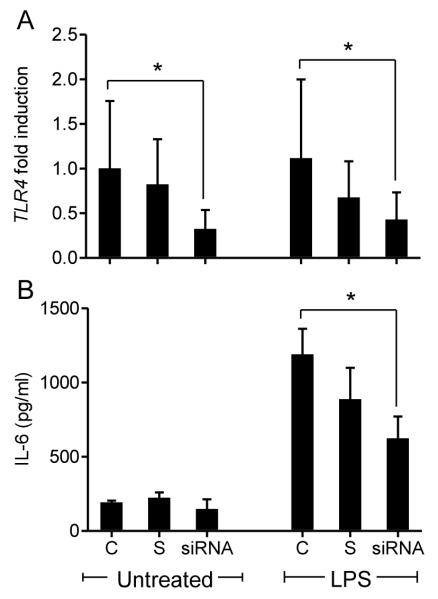
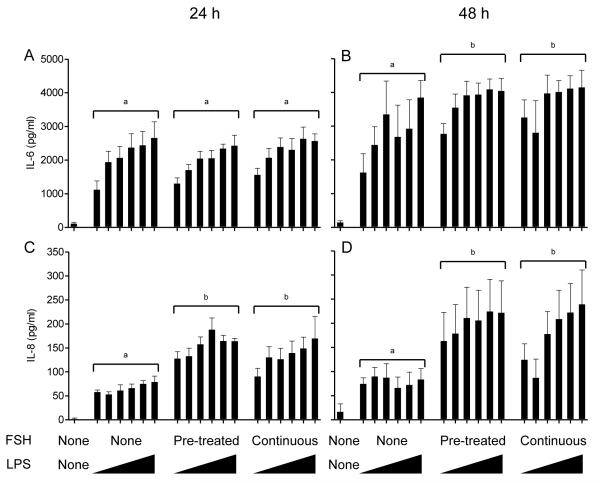
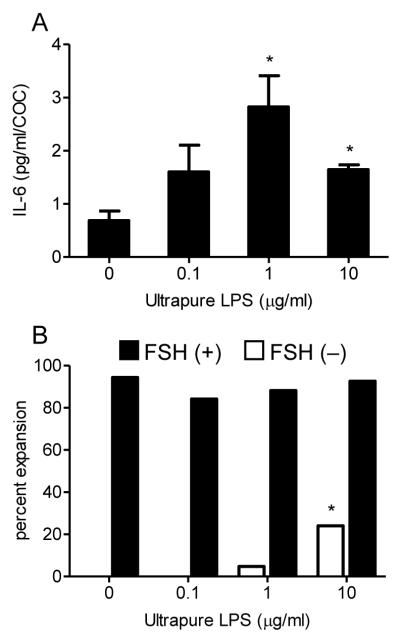
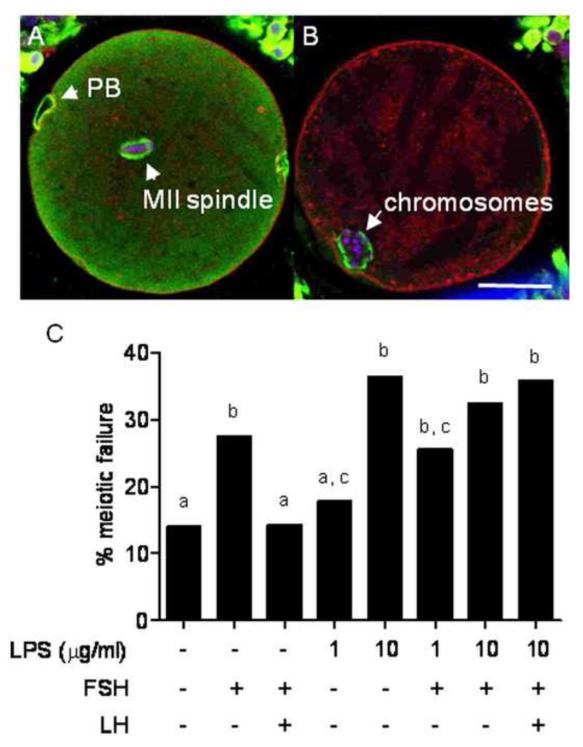
Infections of the reproductive tract or mammary gland with Gram-negative bacteria perturb ovarian function, follicular growth, and fecundity in cattle. We hypothesized that lipopolysaccharide (LPS) from Gram-negative bacteria stimulates an inflammatory response by ovarian granulosa cells that is mediated by Toll-like receptor (TLR) 4. The present study tested the capacity of bovine ovarian granulosa cells to initiate an inflammatory response to pathogen-associated molecular patterns and determined subsequent effects on the in vitro maturation of oocytes. Granulosa cells elicited an inflammatory response to pathogen-associated molecular patterns (LPS, lipoteichoic acid, peptidoglycan, or Pam3CSK4) with accumulation of the cytokine IL-6, and the chemokine IL-8, in a time- and dose-dependent manner. Granulosa cells responded acutely to LPS with rapid phosphorylation of TLR signaling components, p38 and ERK, and increased expression of IL6 and IL8 mRNA, although nuclear translocation of p65 was not evident. Targeting TLR4 with small interfering RNA attenuated granulosa cell accumulation of IL-6 in response to LPS. Endocrine function of granulosa cells is regulated by FSH, but here, FSH also enhanced responsiveness to LPS, increasing IL-6 and IL-8 accumulation. Furthermore, LPS stimulated IL-6 secretion and expansion by cumulus-oocyte complexes and increased rates of meiotic arrest and germinal vesicle breakdown failure. In conclusion, bovine granulosa cells initiate an innate immune response to LPS via the TLR4 pathway, leading to inflammation and to perturbation of meiotic competence.
Figures
References
-
- Sheldon IM, Noakes DE, Rycroft AN, Pfeiffer DU, Dobson H. Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. Reproduction. 2002;123:837–845. - PubMed
-
- Lavon Y, Leitner G, Klipper E, Moallem U, Meidan R, Wolfenson D. Subclinical, chronic intramammary infection lowers steroid concentrations and gene expression in bovine preovulatory follicles. Domest Anim Endocrinol. 2011;40:98–109. - PubMed
-
- Dohmen MJ, Joop K, Sturk A, Bols PE, Lohuis JA. Relationship between intrauterine bacterial contamination, endotoxin levels and the development of endometritis in postpartum cows with dystocia or retained placenta. Theriogenology. 2000;54:1019–1032. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
