

Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet
- PMID: 21990350
- PMCID: PMC3203818
- DOI: 10.1073/pnas.1110655108
Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet
Abstract
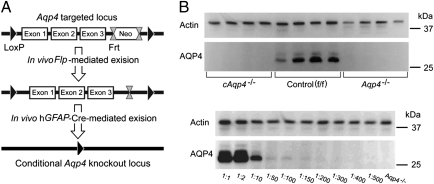
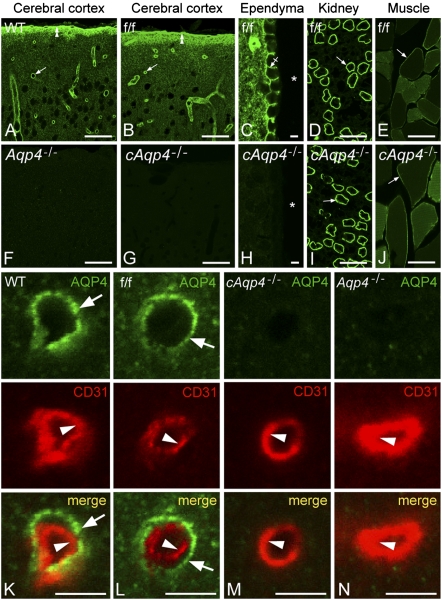
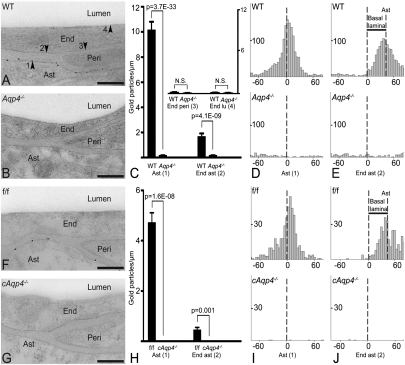
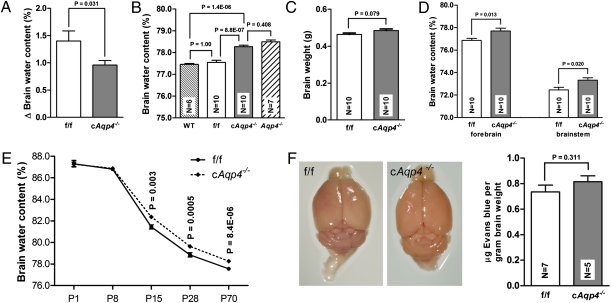
Tissue- and cell-specific deletion of the Aqp4 gene is required to differentiate between the numerous pools of aquaporin-4 (AQP4) water channels. A glial-conditional Aqp4 knockout mouse line was generated to resolve whether astroglial AQP4 controls water exchange across the blood-brain interface. The conditional knockout was driven by the glial fibrillary acidic protein promoter. Brains from conditional Aqp4 knockouts were devoid of AQP4 as assessed by Western blots, ruling out the presence of a significant endothelial pool of AQP4. In agreement, immunofluorescence analysis of cryostate sections and quantitative immunogold analysis of ultrathin sections revealed no AQP4 signals in capillary endothelia. Compared with litter controls, glial-conditional Aqp4 knockout mice showed a 31% reduction in brain water uptake after systemic hypoosmotic stress and a delayed postnatal resorption of brain water. Deletion of astroglial Aqp4 did not affect the barrier function to macromolecules. Our data suggest that the blood-brain barrier (BBB) is more complex than anticipated. Notably, under certain conditions, the astrocyte covering of brain microvessels is rate limiting to water movement.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bechmann I, Galea I, Perry VH. What is the blood-brain barrier (not)? Trends Immunol. 2007;28:5–11. - PubMed
-
- Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006;7:41–53. - PubMed
-
- Manley GT, et al. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat Med. 2000;6:159–163. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
