

Cascades of multisite phosphorylation control Sic1 destruction at the onset of S phase
- PMID: 21993622
- PMCID: PMC3228899
- DOI: 10.1038/nature10560
Cascades of multisite phosphorylation control Sic1 destruction at the onset of S phase
Abstract
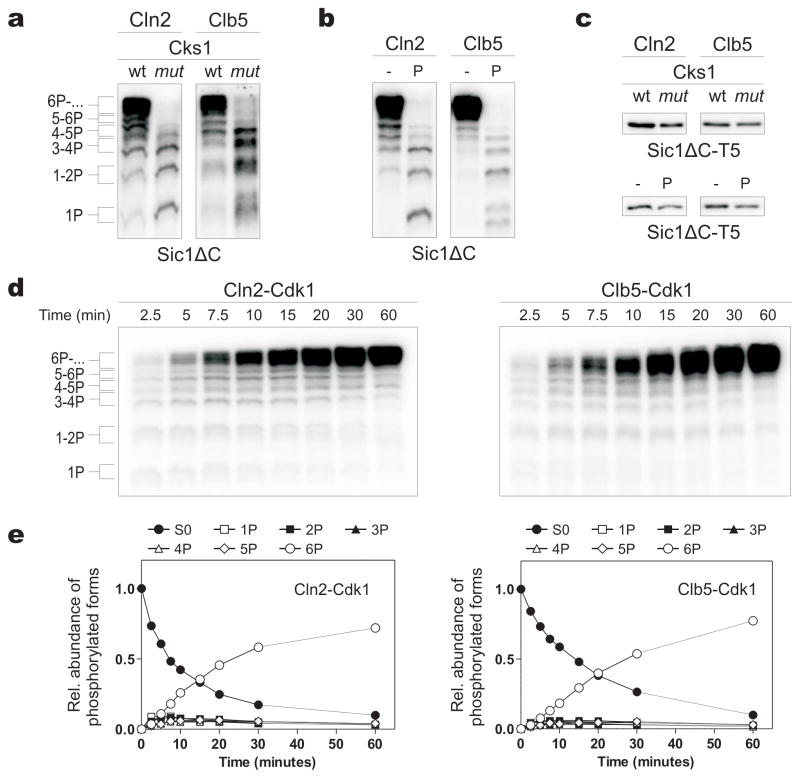
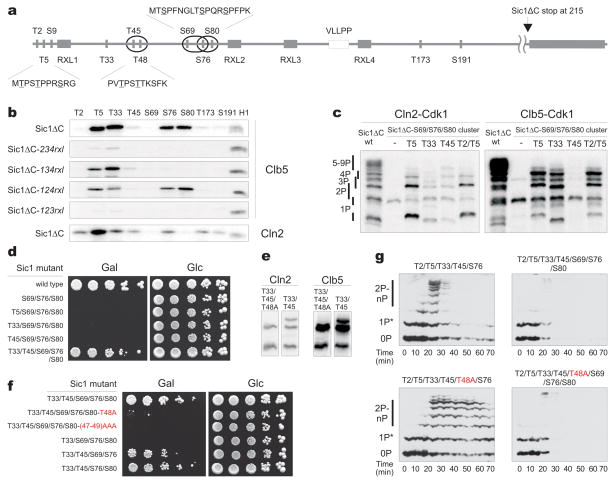
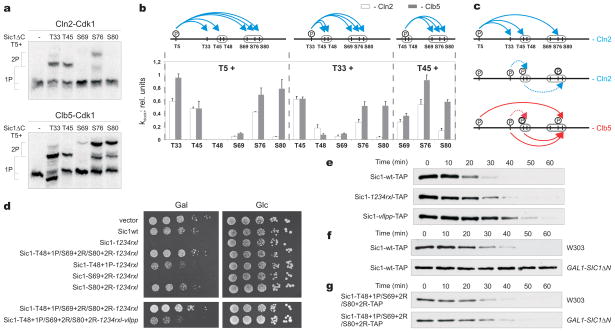
Multisite phosphorylation of proteins has been proposed to transform a graded protein kinase signal into an ultrasensitive switch-like response. Although many multiphosphorylated targets have been identified, the dynamics and sequence of individual phosphorylation events within the multisite phosphorylation process have never been thoroughly studied. In Saccharomyces cerevisiae, the initiation of S phase is thought to be governed by complexes of Cdk1 and Cln cyclins that phosphorylate six or more sites on the Clb5-Cdk1 inhibitor Sic1, directing it to SCF-mediated destruction. The resulting Sic1-free Clb5-Cdk1 complex triggers S phase. Here, we demonstrate that Sic1 destruction depends on a more complex process in which both Cln2-Cdk1 and Clb5-Cdk1 act in processive multiphosphorylation cascades leading to the phosphorylation of a small number of specific phosphodegrons. The routes of these phosphorylation cascades are shaped by precisely oriented docking interactions mediated by cyclin-specific docking motifs in Sic1 and by Cks1, the phospho-adaptor subunit of Cdk1. Our results indicate that Clb5-Cdk1-dependent phosphorylation generates positive feedback that is required for switch-like Sic1 destruction. Our evidence for a docking network within clusters of phosphorylation sites uncovers a new level of complexity in Cdk1-dependent regulation of cell cycle transitions, and has general implications for the regulation of cellular processes by multisite phosphorylation.
Figures
References
-
- Nash P, et al. Multisite phosphorylation of a CDK inhibitor sets a threshold for the onset of DNA replication. Nature. 2001;414:514–521. - PubMed
-
- Kim SY, Ferrell JE., Jr Substrate competition as a source of ultrasensitivity in the inactivation of Wee1. Cell. 2007;128:1133–1145. - PubMed
-
- Tang X, et al. Suprafacial orientation of the SCFCdc4 dimer accommodates multiple geometries for substrate ubiquitination. Cell. 2007;129:1165–1176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
