

Vocal control by the common marmoset in the presence of interfering noise
- PMID: 21993791
- PMCID: PMC3192021
- DOI: 10.1242/jeb.056101
Vocal control by the common marmoset in the presence of interfering noise
Abstract
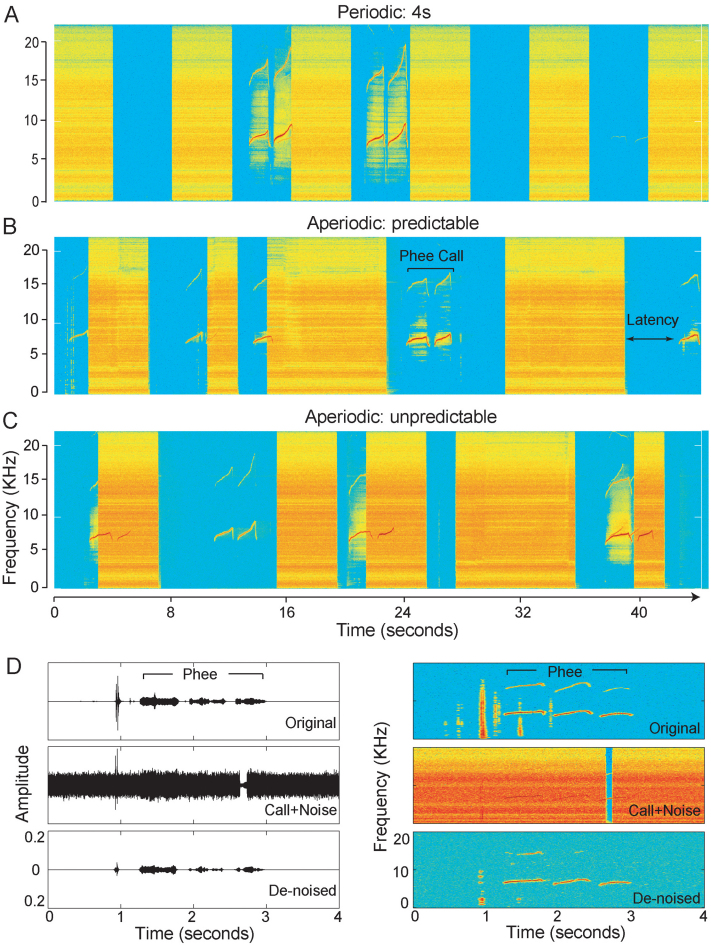
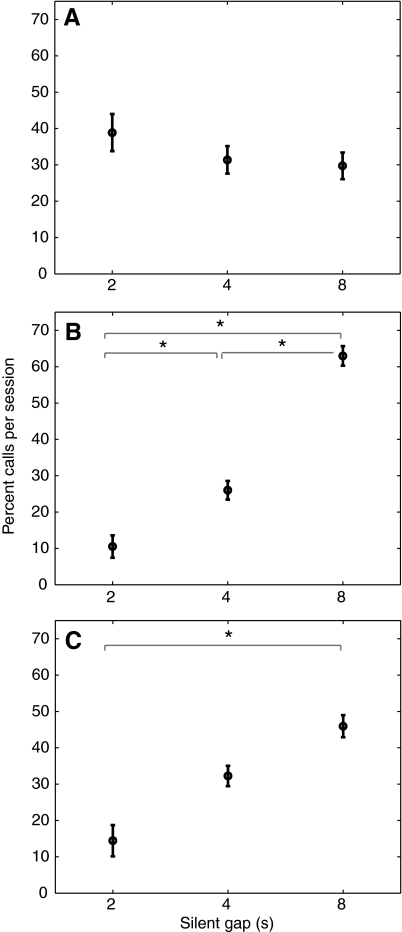
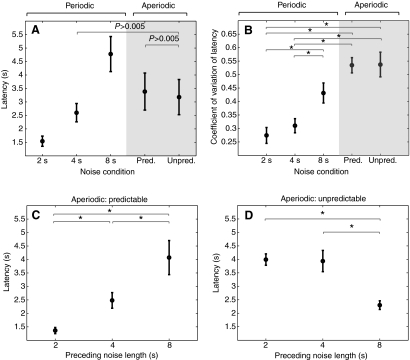
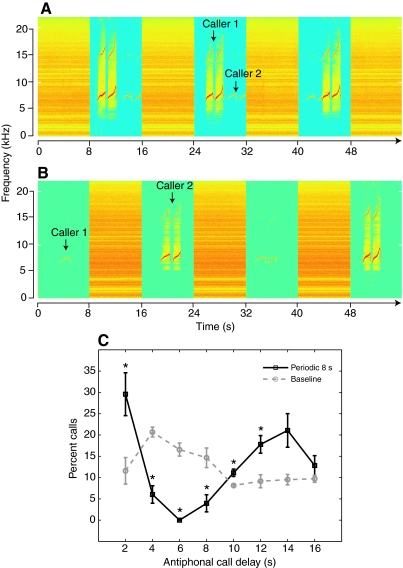
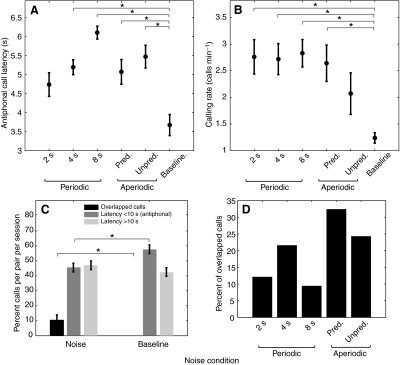
The natural environment is inherently noisy with acoustic interferences. It is, therefore, beneficial for a species to modify its vocal production to effectively communicate in the presence of interfering noises. Non-human primates have been traditionally considered to possess limited voluntary vocal control, but little is known about their ability to modify vocal behavior when encountering interfering noises. Here we tested the ability of the common marmoset (Callithrix jacchus) to control the initiation of vocalizations and maintain vocal interactions between pairs in an acoustic environment in which the length and predictability (periodic or random aperiodic occurrences) of interfering noise bursts were varied. Despite the presence of interfering noise, the marmosets continued to engage in antiphonal calling behavior. Results showed that the overwhelming majority of calls were initiated during silence gaps even when the length of the silence gap following each noise burst was unpredictable. During the periodic noise conditions, as the length of the silence gap decreased, the latency between the end of noise burst and call onset decreased significantly. In contrast, when presented with aperiodic noise bursts, the marmosets chose to call predominantly during long (4 and 8 s) over short (2 s) silence gaps. In the 8 s periodic noise conditions, a marmoset pair either initiated both calls of an antiphonal exchange within the same silence gap or exchanged calls in two consecutive silence gaps. Our findings provide compelling evidence that common marmosets are capable of modifying their vocal production according to the dynamics of their acoustic environment during vocal communication.
Figures
References
-
- Bezerra B. M., Souto A. (2008). Structure and usage of the vocal repertoire of Callithrix jacchus. Int. J. Primatol. 29, 671-701
-
- Brumm H. (2006). Signalling through acoustic windows: nightingales avoid interspecific competition by short-term adjustment of song timing. J. Comp. Physiol. A 192, 1279-1285 - PubMed
-
- Brumm H., Slabbekoorn H. (2005). Acoustic communication in noise. Adv. Stud. Behav. 35, 151-209
-
- Brumm H., Voss K., Kollmer I., Todt D. (2004). Acoustic communication in noise: regulation of call characteristics in a New World monkey. J. Exp. Biol. 207, 443-448 - PubMed
-
- Brumm H., Schmidt R., Schrader L. (2009). Noise-dependent vocal plasticity in domestic fowl. Anim. Behav. 78, 741-746
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
