

Hepatitis C virus activates interleukin-1β via caspase-1-inflammasome complex
- PMID: 21994322
- PMCID: PMC3352344
- DOI: 10.1099/vir.0.034033-0
Hepatitis C virus activates interleukin-1β via caspase-1-inflammasome complex
Retraction in
-
Retraction notice: Hepatitis C virus activates interleukin-1β via caspase-1-inflammasome complex.J Gen Virol. 2019 Dec;100(12):1714. doi: 10.1099/jgv.0.001339. J Gen Virol. 2019. PMID: 31647405 Free PMC article. No abstract available.
Expression of concern in
-
Expression of concern: Hepatitis C virus activates interleukin-1β via caspase-1-inflammasome complex.J Gen Virol. 2019 Sep;100(9):1342. doi: 10.1099/jgv.0.001313. Epub 2019 Aug 23. J Gen Virol. 2019. PMID: 31441743 Free PMC article. No abstract available.
Abstract
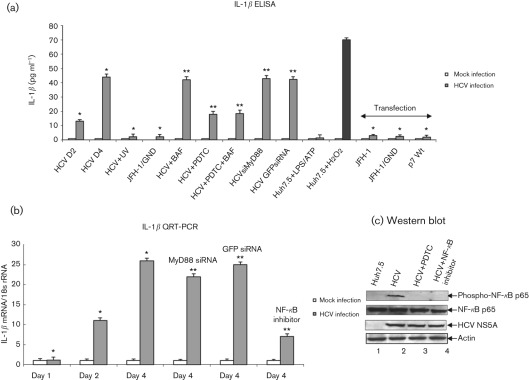
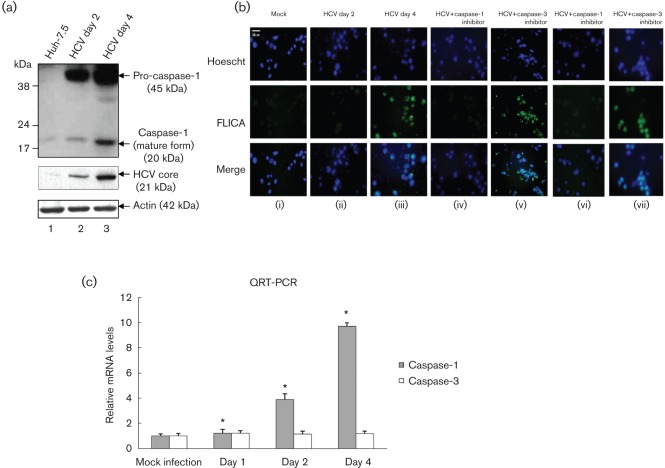
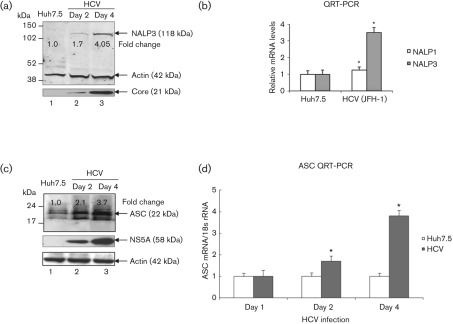
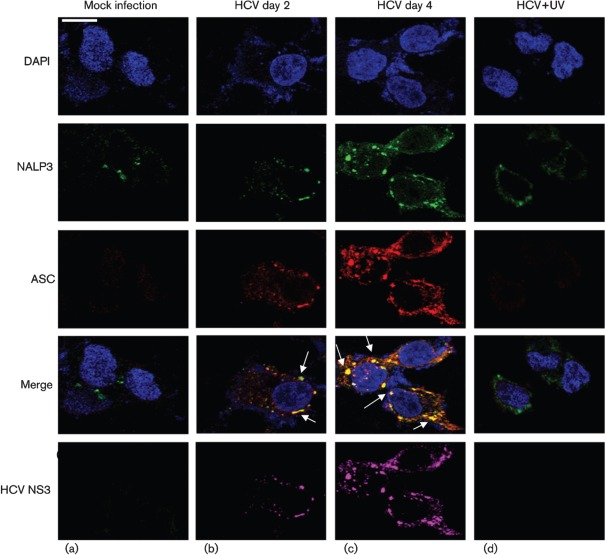
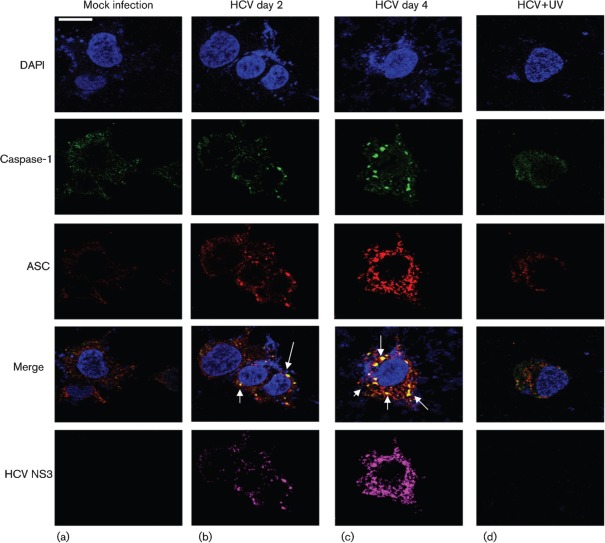
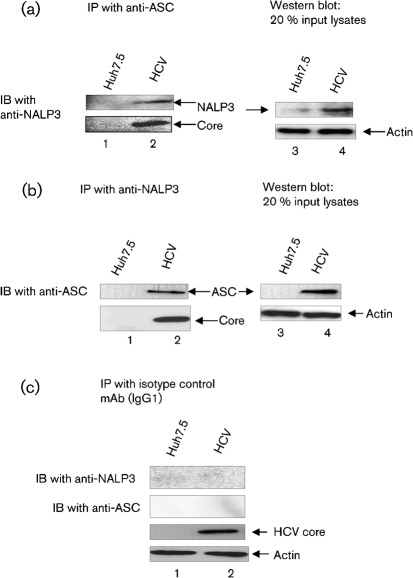
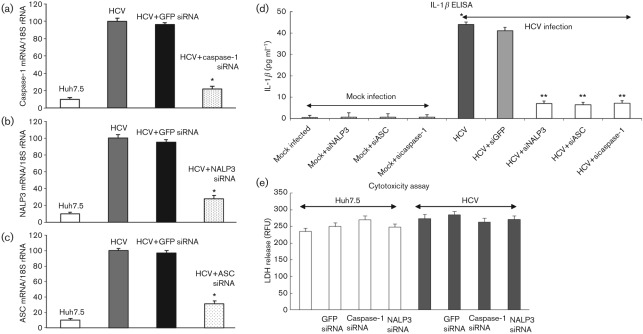
Interleukin-1β (IL-1β) is a potent pro-inflammatory cytokine involved in the pathogenesis of HCV, but the sensors and underlying mechanisms that facilitate HCV-induced IL-1β proteolytic activation and secretion remains unclear. In this study, we have identified a signalling pathway leading to IL-1β activation and secretion in response to HCV infection. Previous studies have shown the induction and secretion of IL-1β through the inflammasome complex in macrophages/monocytes. Here, we report for the first time the induction and assembly of the NALP3-inflammasome complex in human hepatoma cells infected with HCV (JFH-1). We demonstrate that activation of IL-1β in HCV-infected cells involves the proteolytic processing of pro-caspase-1 into mature caspase-1 in a multiprotein inflammasome complex. Next, we demonstrate that HCV is sensed by NALP3 protein, which recruits the adaptor protein ASC for the assembly of the inflammasome complex. Using a small interfering RNA approach, we further show that components of the inflammasome complex are involved in the activation of IL-1β in HCV-infected cells. Our study also demonstrates the role of reactive oxygen species in HCV-induced IL-1β secretion. Collectively, these observations provide an insight into the mechanism of IL-1β processing and secretion, which is likely to provide novel strategies for targeting the viral or cellular determinants to arrest the progression of liver disease associated with chronic HCV infection.
Figures
References
-
- Allen I. C., Scull M. A., Moore C. B., Holl E. K., McElvania-TeKippe E., Taxman D. J., Guthrie E. H., Pickles R. J., Ting J. P. (2009). The NLRP3 inflammasome mediates in vivo innate immunity to influenza A virus through recognition of viral RNA. Immunity 30, 556–565 10.1016/j.immuni.2009.02.005 - DOI - PMC - PubMed
-
- Bartenschlager R., Lohmann V. (2000). Replication of hepatitis C virus. J Gen Virol 81, 1631–1648 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous