

Receptor agonism and antagonism of dietary bitter compounds
- PMID: 21994393
- PMCID: PMC6703385
- DOI: 10.1523/JNEUROSCI.2923-11.2011
Receptor agonism and antagonism of dietary bitter compounds
Abstract
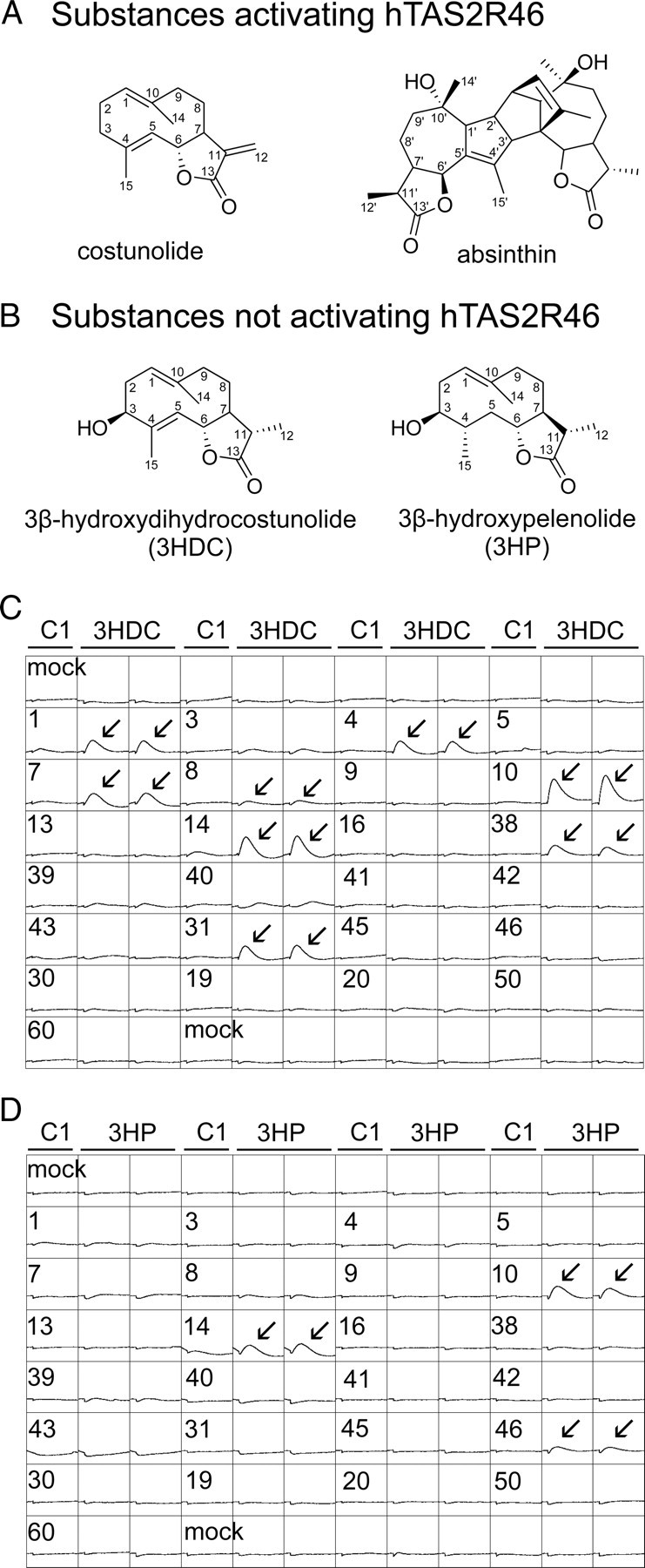
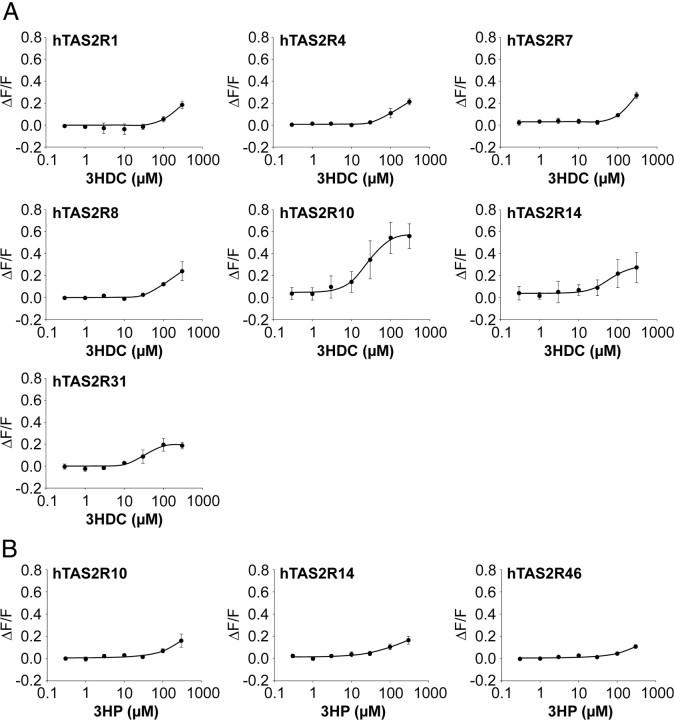
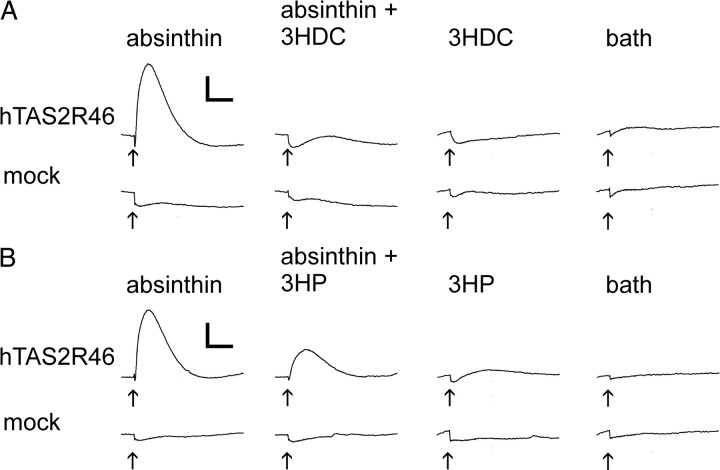
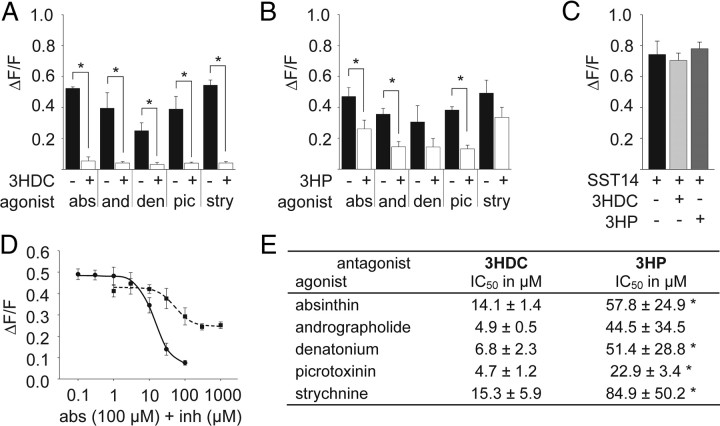
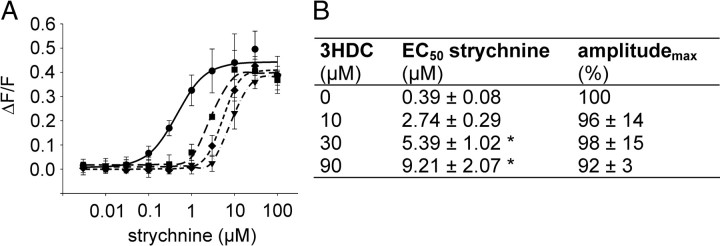
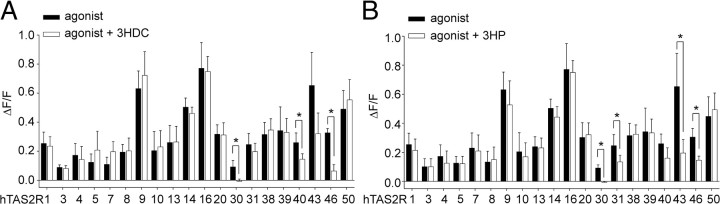
Food contains complex blends of structurally diverse bitter compounds that trigger bitterness through activation of one or more of the ∼25 human TAS2 bitter taste receptors. It remains unsolved, however, whether the perceived bitterness of binary bitter-compound mixtures can be considered an additive function of all bitter-inducing chemicals in the mouth, suggesting that little mutual interaction takes place among bitter substances or if mixture suppression and synergism occurs. Here we report on two natural sesquiterpene lactones from edible plants, which stimulate distinct sets of hTAS2Rs in transfected cells. Both chemicals also robustly inhibit different but overlapping subsets of agonist-activated hTAS2Rs. These findings demonstrate that mixtures of bitter compounds, because they normally occur in human foodstuff, likely elicit bitter perception in a complex and not in a merely additive manner. An unexpected implication of this discovery is that, during evolution, the naturally occurring bitter taste receptor antagonists have shaped some of the pharmacological properties of the receptors, such as overlapping recognition profiles and breadth of tuning.
Figures
References
-
- Ache BW, Young JM. Olfaction: diverse species, conserved principles. Neuron. 2005;48:417–430. - PubMed
-
- Akhmedov IS, Kasymov SZ, Sidyakin GP. Artabin: a new lactone from Artemisia absinthium. Chem Nat Compounds. 1970;6:634.
-
- Ammon C, Schäfer J, Kreuzer OJ, Meyerhof W. Presence of a plasma membrane targeting sequence in the amino-terminal region of the rat somatostatin receptor 3. Arch Physiol Biochem. 2002;110:137–145. - PubMed
-
- Ayya N, Lawless HT. Quantitative and qualitative evaluation of high-intensity sweeteners and sweetener mixtures. Chem Senses. 1992;17:245–259.
-
- Bartoshuk LM. Taste mixtures: is mixture suppression related to compression? Physiol Behav. 1975;14:643–649. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases