

Defective Interfering RNAs: Foes of Viruses and Friends of Virologists
- PMID: 21994575
- PMCID: PMC3185524
- DOI: 10.3390/v1030895
Defective Interfering RNAs: Foes of Viruses and Friends of Virologists
Abstract
Defective interfering (DI) RNAs are subviral RNAs produced during multiplication of RNA viruses by the error-prone viral replicase. DI-RNAs are parasitic RNAs that are derived from and associated with the parent virus, taking advantage of viral-coded protein factors for their multiplication. Recent advances in the field of DI RNA biology has led to a greater understanding about generation and evolution of DI-RNAs as well as the mechanism of symptom attenuation. Moreover, DI-RNAs are versatile tools in the hands of virologists and are used as less complex surrogate templates to understand the biology of their helper viruses. The ease of their genetic manipulation has resulted in rapid discoveries on cis-acting RNA replication elements required for replication and recombination. DI-RNAs have been further exploited to discover host factors that modulate Tomato bushy stunt virus replication, as well as viral RNA recombination. This review discusses the current models on generation and evolution of DI-RNAs, the roles of viral and host factors in DI-RNA replication, and the mechanisms of disease attenuation.
Keywords: RNA structure; RNA virus; host factors; recombination; replication.
Figures
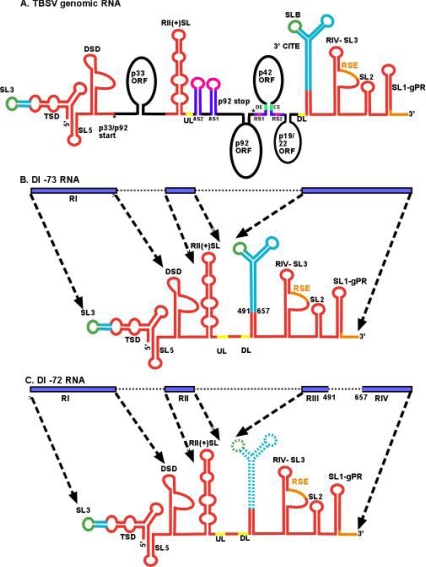
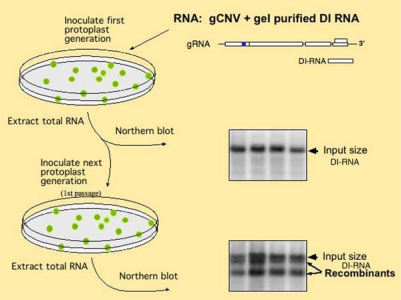
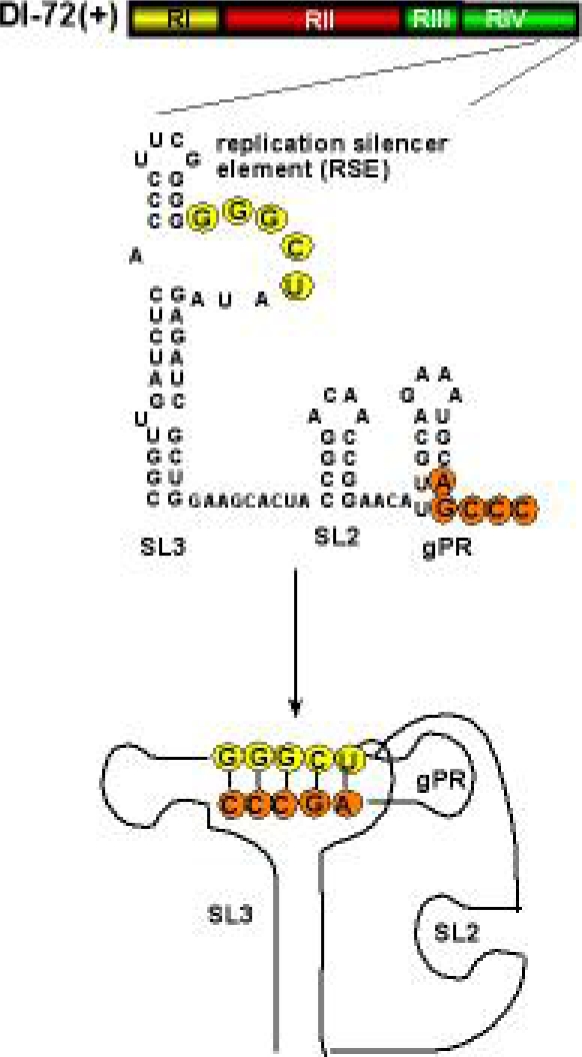
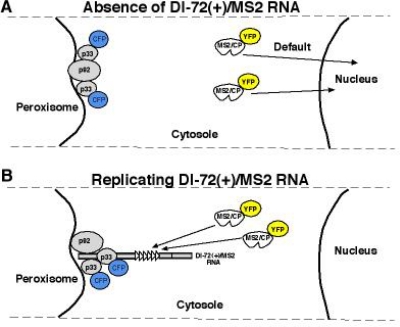
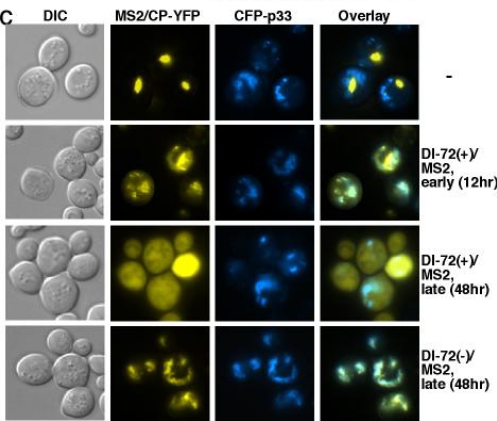
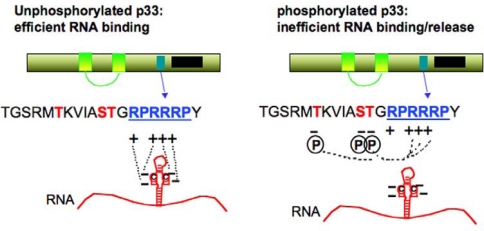
References
-
- Huang AS. Defective interfering viruses. Annu Rev Microbiol. 1973;27:101–117. - PubMed
-
- Simon AE, Roossinck MJ, Havelda Z. Plant virus satellite and defective interfering RNAs: new paradigms for a new century. Annu Rev Phytopathol. 2004;42:415–437. - PubMed
-
- Perrault J. Origin and replication of defective interfering particles. Curr Top Microbiol Immunol. 1981;93:151–207. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
