

Recent advances in hepatitis C virus cell entry
- PMID: 21994653
- PMCID: PMC3185649
- DOI: 10.3390/v2030692
Recent advances in hepatitis C virus cell entry
Abstract
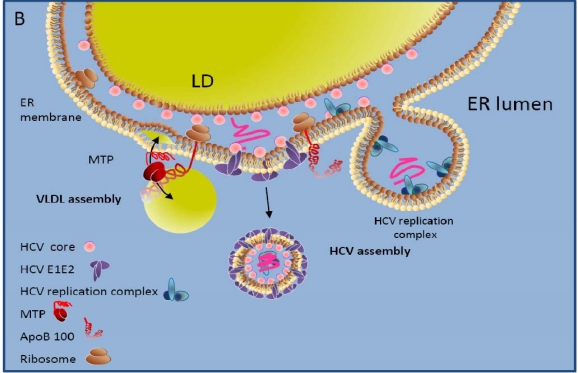
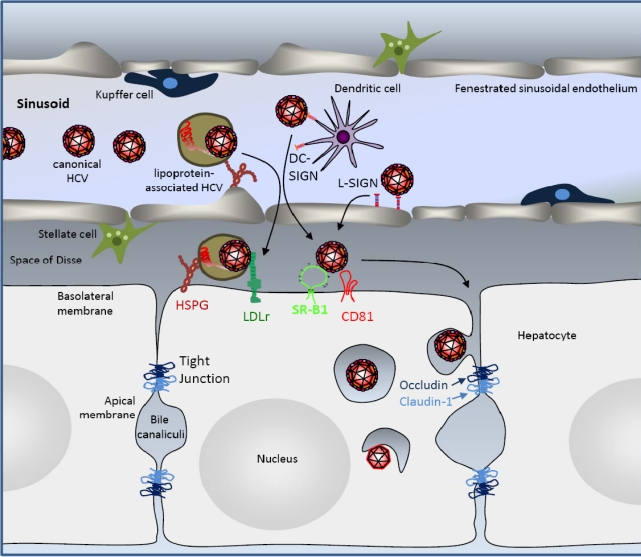
More than 170 million patients worldwide are chronically infected with hepatitis C virus (HCV). Prevalence rates range from 0.5% in Northern European countries to 28% in some areas of Egypt. HCV is hepatotropic, and in many countries chronic hepatitis C is a leading cause of liver disease including fibrosis, cirrhosis and hepatocellular carcinoma. HCV persists in 50-85% of infected patients, and once chronic infection is established, spontaneous clearance is rare. HCV is a member of the Flaviviridae family, in which it forms its own genus. Many lines of evidence suggest that the HCV life cycle displays many differences to that of other Flaviviridae family members. Some of these differences may be due to the close interaction of HCV with its host's lipid and particular triglyceride metabolism in the liver, which may explain why the virus can be found in association with lipoproteins in serum of infected patients. This review focuses on the molecular events underlying the HCV cell entry process and the respective roles of cellular co-factors that have been implied in these events. These include, among others, the lipoprotein receptors low density lipoprotein receptor and scavenger receptor BI, the tight junction factors occludin and claudin-1 as well as the tetraspanin CD81. We discuss the roles of these cellular factors in HCV cell entry and how association of HCV with lipoproteins may modulate the cell entry process.
Keywords: cell entry; hepatitis C virus.
Figures

References
-
- Moradpour D, Penin F, Rice CM. Replication of hepatitis C virus. Nat Rev Microbiol. 2007;5:453–463. - PubMed
-
- Lindenbach BD, Thiel HJ, Rice CM. Flaviviridae: The Viruses and Their Replication. In: Knipe DM, Howley PM, editors. Fields Virology. 5th ed. Vol. 1. Lippincott Williams & Wilkins; PA, USA: 2007. pp. 1101–1152.
-
- Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, Zayas M, Bartenschlager R, Wakita T, Hijikata M, Shimotohno K. The lipid droplet is an important organelle for hepatitis C virus production. Nat Cell Biol. 2007;9:1089–1097. - PubMed
LinkOut - more resources
Full Text Sources
