

Efficient sensing of avian influenza viruses by porcine plasmacytoid dendritic cells
- PMID: 21994734
- PMCID: PMC3185703
- DOI: 10.3390/v3040312
Efficient sensing of avian influenza viruses by porcine plasmacytoid dendritic cells
Abstract
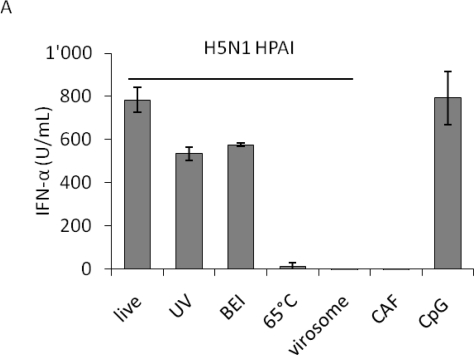
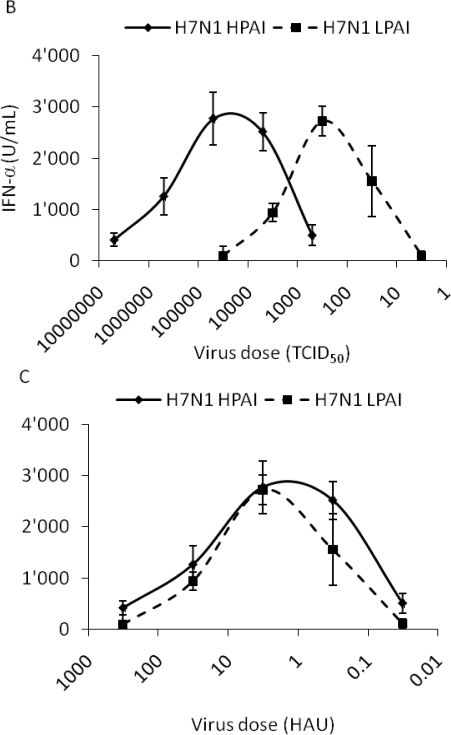
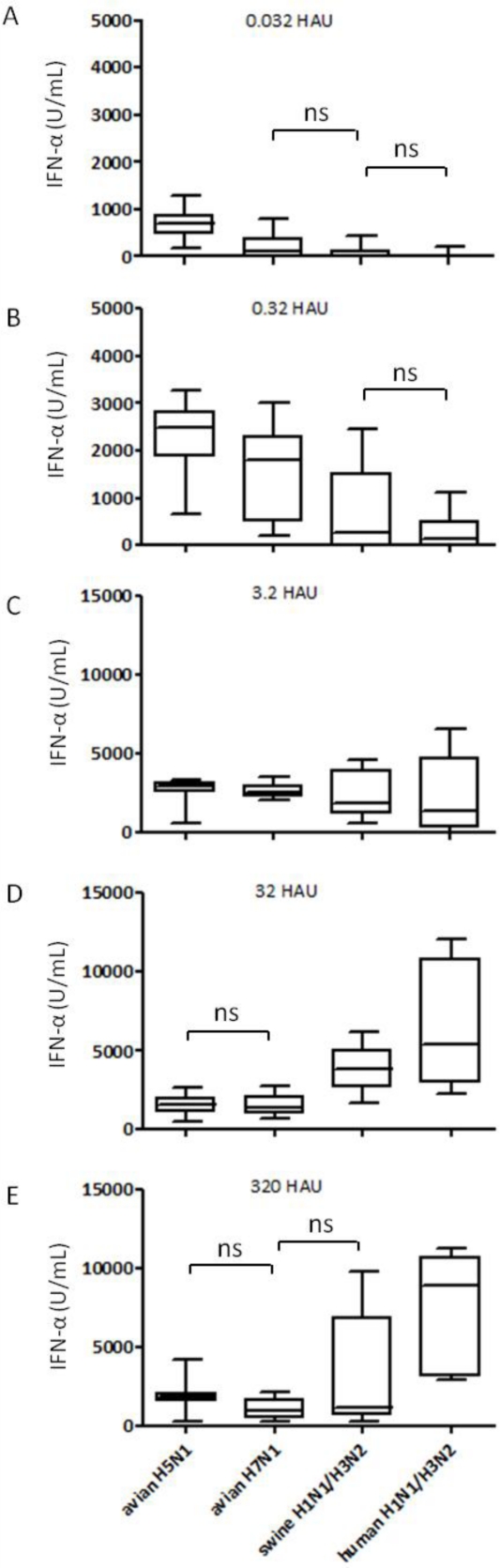
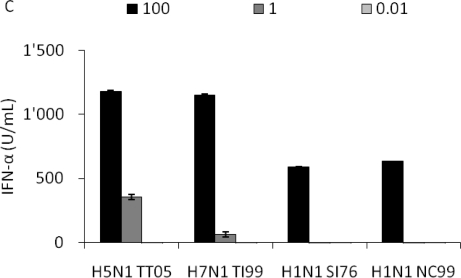
H5N1 influenza A virus (IAV) infections in human remain rare events but have been associated with severe disease and a higher mortality rate compared to infections with seasonal strains. An excessive release of pro-inflammatory cytokine together with a greater virus dissemination potential have been proposed to explain the high virulence observed in human and other mammalian and avian species. Among the cells involved in the cytokine storm, plasmacytoid dendritic cells (pDC) could play an important role considering their unique capacity to secrete massive amounts of type I interferon (IFN). Considering the role of IFN as a major component of antiviral responses as well as in priming inflammatory responses, we aimed to characterize the induction of IFN-α release upon infection with IAV originating from various avian and mammalian species in a comparative way. In our porcine pDC model, we showed that the viral components triggering IFN responses related to the ability to hemagglutinate, although virosomes devoid of viral RNA were non-stimulatory. Heat-treatment at 65 °C but not chemical inactivation destroyed the ability of IAV to stimulate pDC. All IAV tested induced IFN-α but at different levels and showed different dose-dependencies. H5 and H7 subtypes, in particular H5N1, stimulated pDC at lower doses when compared to mammalian IAV. At high viral doses, IFN-α levels reached by some mammalian IAV surpassed those induced by avian isolates. Although sialic acid-dependent entry was demonstrated, the α-2,3 or α-2,6 binding specificity alone did not explain the differences observed. Furthermore, we were unable to identify a clear role of the hemagglutinin, as the IFN-α doses-response profiles did not clearly differ when viruses with all genes of identical avian origin but different HA were compared. This was found with IAV bearing an HA derived from either a low, a high pathogenic H5N1, or a human H3. Stimulation of pDC was associated with pDC depletion within the cultures. Taken together and considering the efficient sensing of H5N1 at low dose, pDC on one side may play a role in the cytokine storm observed during severe disease, on the other hand could participate in early antiviral responses limiting virus replication.
Keywords: cytokine storm; influenza A virus; interferon; plasmacytoid dendritic cells.
Figures
References
-
- Beigel JH, Farrar J, Han AM, Hayden FG, Hyer R, de Jong MD, Lochindarat S, Nguyen TK, Nguyen TH, Tran TH, et al. Avian influenza A (H5N1) infection in humans. N. Engl. J. Med. 2005;353:1374–1385. - PubMed
-
- Cheung CY, Poon LL, Lau AS, Luk W, Lau YL, Shortridge KF, Gordon S, Guan Y, Peiris JS. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: A mechanism for the unusual severity of human disease? Lancet. 2002;360:1831–1837. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
