

The emerging functions of septins in metazoans
- PMID: 21997296
- PMCID: PMC3207108
- DOI: 10.1038/embor.2011.193
The emerging functions of septins in metazoans
Abstract
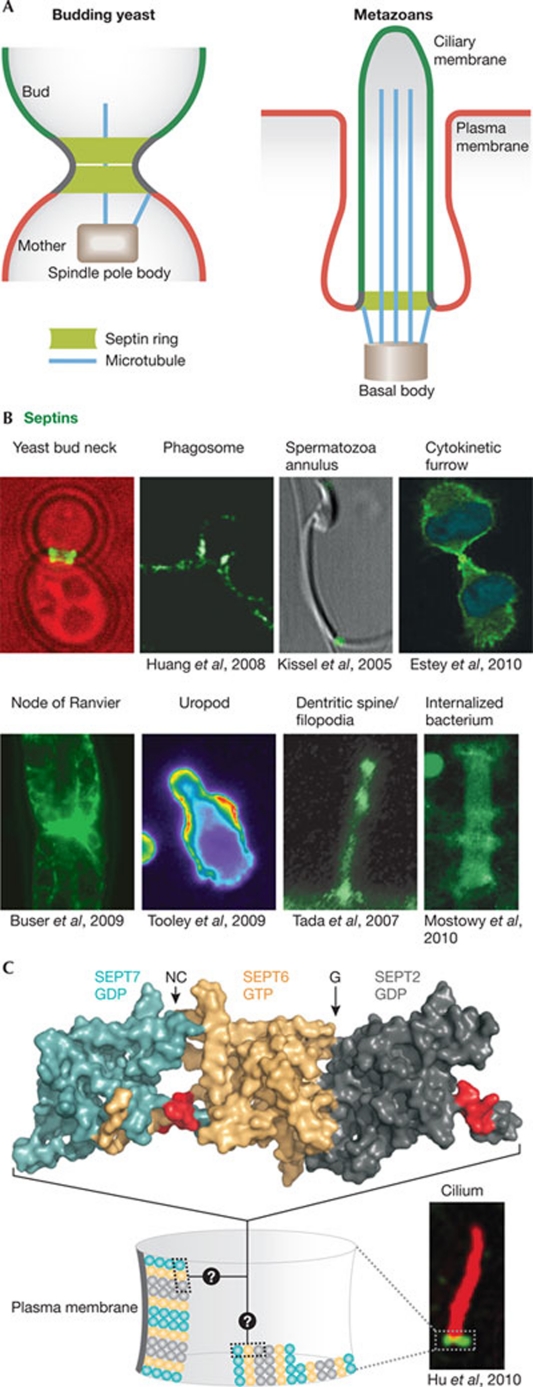
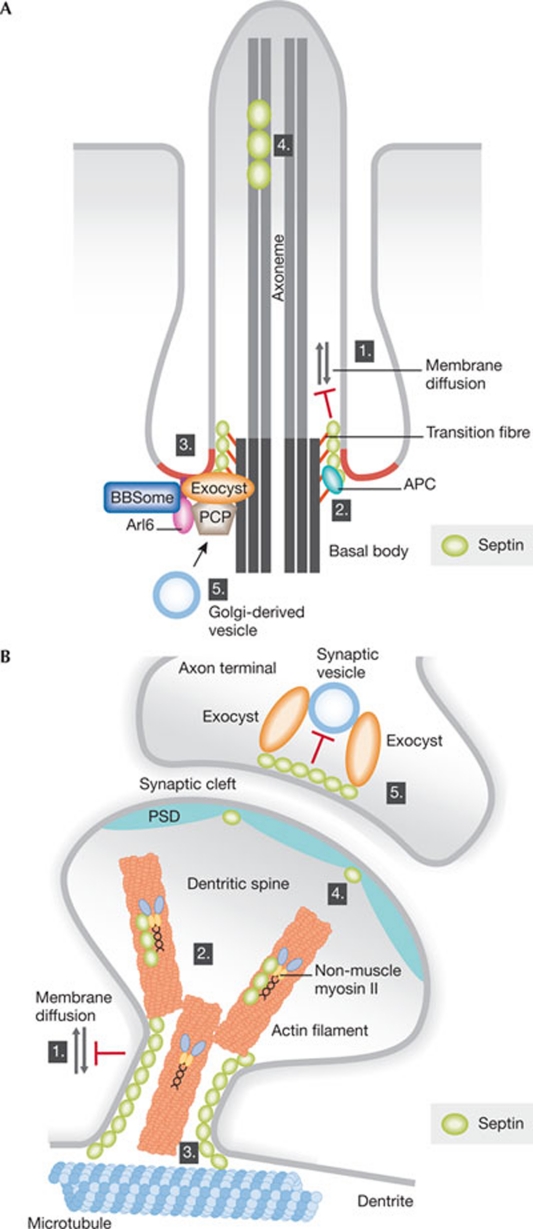
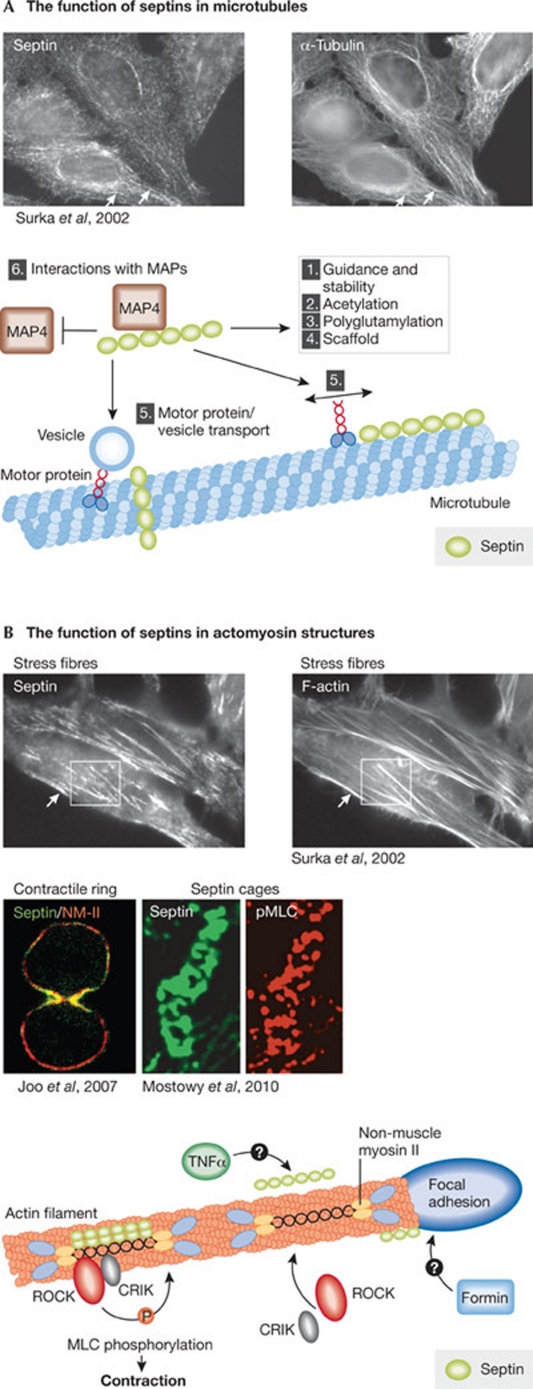
Septins form a subfamily of highly related GTP-binding proteins conserved from eukaryotic protists to mammals. In most cases, septins function in close association with cell membranes and the actin and microtubule cytoskeleton to regulate a wide variety of key cellular processes. Further underscoring their importance, septin abnormalities are associated with several human diseases. Remarkably, septins have the ability to polymerize into assemblies of different sizes in vitro and in vivo. In cells, these structures act in the formation of diffusion barriers and scaffolds that maintain subcellular polarity. Here, we focus on the emerging roles of vertebrate septins in ciliogenesis, neurogenesis, tumorigenesis and host-pathogen interactions, and discuss whether unifying themes underlie the molecular function of septins in health and disease.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Barral Y, Mermall V, Mooseker MS, Syder M (2000) Compartmentalization of the cell cortex by septins is required for the maintenance of cell polarity in yeast. Mol Cell 5: 841–851 - PubMed
-
- Beites CL, Xie H, Bowser R, Trimble WS (1999) The septin CDCrel-1 binds syntaxin and inhibits exocytosis. Nat Neurosci 2: 434–439 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
