

Hypoxic regulation of pulmonary vascular smooth muscle cyclic guanosine monophosphate-dependent kinase by the ubiquitin conjugating system
- PMID: 21997485
- PMCID: PMC3326432
- DOI: 10.1165/rcmb.2011-0165OC
Hypoxic regulation of pulmonary vascular smooth muscle cyclic guanosine monophosphate-dependent kinase by the ubiquitin conjugating system
Abstract
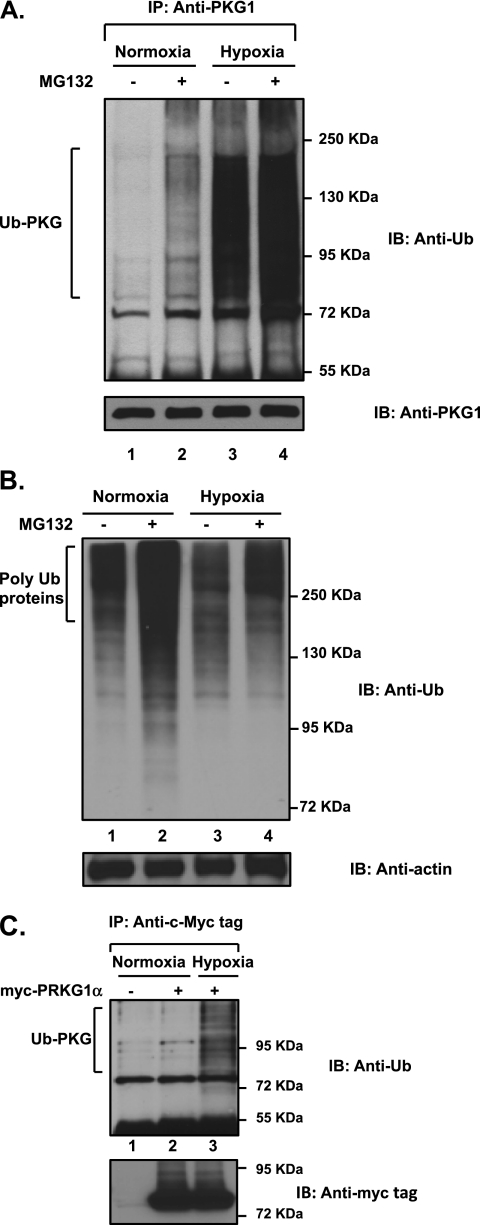
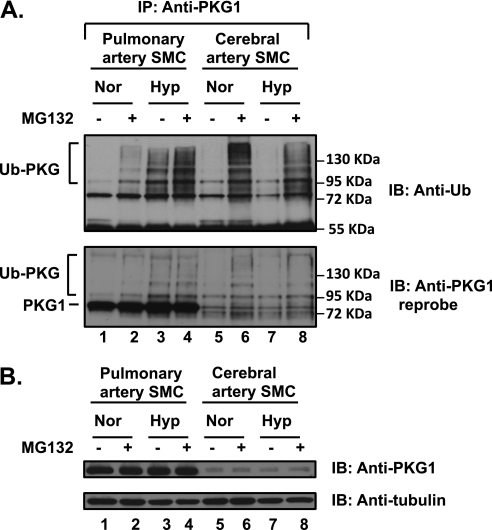
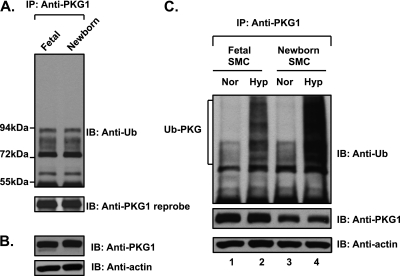
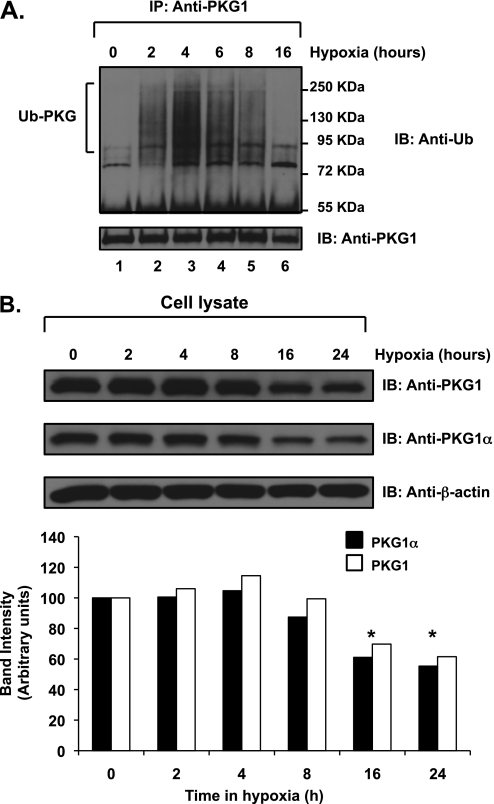
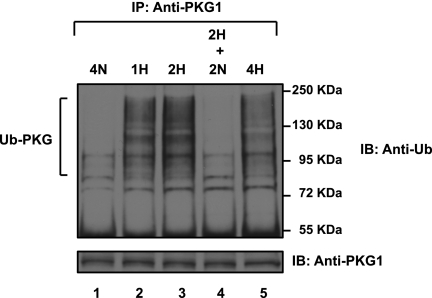
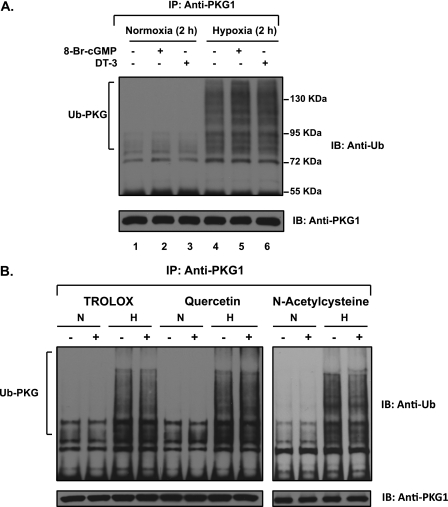
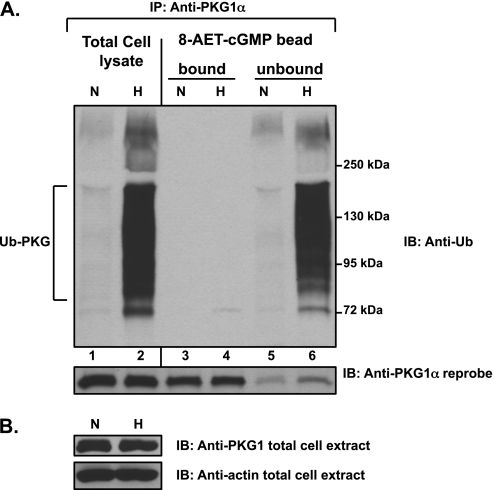
We previously reported that hypoxia attenuates nitric oxide-cyclic guanosine monophosphate (NO-cGMP)-mediated fetal pulmonary vessel relaxation by inhibiting cGMP-dependent protein kinase 1 (PKG1) activity, but not all the mechanisms by which acute hypoxia inhibits PKG1 activity have been delineated. Here we demonstrate for the first time, to the best of our knowledge, that acute hypoxia induces an accumulation of ubiquitinated PKG1 in ovine fetal and newborn pulmonary artery smooth muscle cells. Such a modification was not evident in ovine fetal systemic (cerebral) artery smooth muscle cells. The accumulation of polyubiquitinated PKG1 observed after 4 hours of hypoxia was affected neither by the activation of PKG1 kinase activity with the cell-permeable cGMP analogue 8-bromo-cGMP, nor by its inhibition with DT-3 in fetal pulmonary artery smooth muscle cells. Ubiquitinated PKG1α was unable to bind the cGMP analogue 8-(2-aminoethyl)thioguanosine-3',5' (AET)-cGMP, a ligand for the unmodified protein. Inhibition of the proteasomal complex with MG132 led to the accumulation of polyubiquitinated PKG1 in normoxia, indicating the involvement of the ubiquitin-26S proteasomal system in degradation and clearance of this protein under normoxic conditions. The ubiquitinated PKG1 under hypoxic conditions, however, was not predominantly targeted for proteasomal degradation. Importantly, reoxygenation reversed the acute hypoxia-induced accumulation of ubiquitinated PKG1. Our results suggest that the PKG1 ubiquitination induced by acute hypoxia plays a unique role in the regulation of the pulmonary vascular smooth muscle cell vasoreactivity and relaxation mediated by the NO-cGMP-PKG1 pathway.
Figures
References
-
- Hofmann F, Bernhard D, Lukowski R, Weinmeister P. cGMP regulated protein kinases (CGK). Handb Exp Pharmacol 2009;191:137–162 - PubMed
-
- Schlossmann J, Desch M. CGK substrates. Handb Exp Pharmacol 2009;191:163–193 - PubMed
-
- Hofmann F, Feil R, Kleppisch T, Schlossmann J. Function of cGMP-dependent protein kinases as revealed by gene deletion. Physiol Rev 2006;86:1–23 - PubMed
-
- Surks HK, Mochizuki N, Kasai Y, Georgescu SP, Tang KM, Ito M, Lincoln TM, Mendelsohn ME. Regulation of myosin phosphatase by a specific interaction with cGMP-dependent protein kinase Ialpha. Science 1999;286:1583–1587 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
