

Impaired Coenzyme A metabolism affects histone and tubulin acetylation in Drosophila and human cell models of pantothenate kinase associated neurodegeneration
- PMID: 21998097
- PMCID: PMC3377114
- DOI: 10.1002/emmm.201100180
Impaired Coenzyme A metabolism affects histone and tubulin acetylation in Drosophila and human cell models of pantothenate kinase associated neurodegeneration
Abstract
Pantothenate kinase-associated neurodegeneration (PKAN is a neurodegenerative disease with unresolved pathophysiology. Previously, we observed reduced Coenzyme A levels in a Drosophila model for PKAN. Coenzyme A is required for acetyl-Coenzyme A synthesis and acyl groups from the latter are transferred to lysine residues of proteins, in a reaction regulated by acetyltransferases. The tight balance between acetyltransferases and their antagonistic counterparts histone deacetylases is a well-known determining factor for the acetylation status of proteins. However, the influence of Coenzyme A levels on protein acetylation is unknown. Here we investigate whether decreased levels of the central metabolite Coenzyme A induce alterations in protein acetylation and whether this correlates with specific phenotypes of PKAN models. We show that in various organisms proper Coenzyme A metabolism is required for maintenance of histone- and tubulin acetylation, and decreased acetylation of these proteins is associated with an impaired DNA damage response, decreased locomotor function and decreased survival. Decreased protein acetylation and the concurrent phenotypes are partly rescued by pantethine and HDAC inhibitors, suggesting possible directions for future PKAN therapy development.
Copyright © 2011 EMBO Molecular Medicine.
Figures
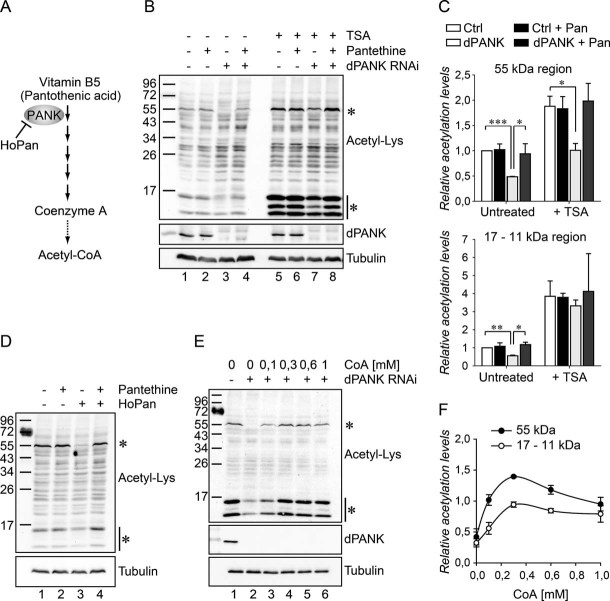
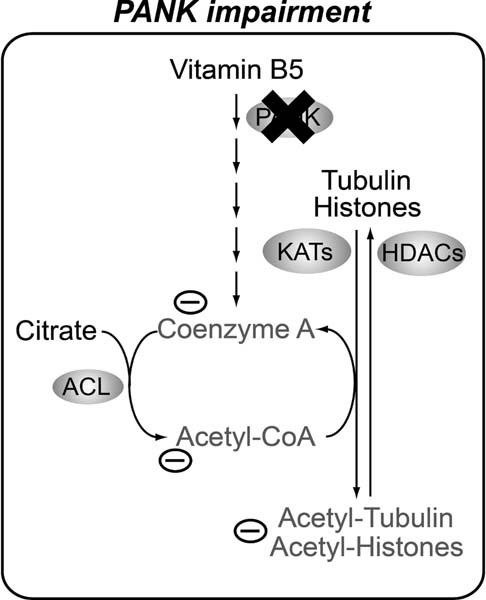
The de novo biosynthesis route of Coenzyme A: Vitamin B5 is converted in several steps into CoA. PANK (referred to as dPANK/Fbl in Drosophila) is required for the first conversion step. HoPan is a potent chemical inhibitor of PANK enzymatic activity.
Whole cell lysates of control S2 cells and dPANK/Fbl-depleted cells (by RNAi) were used to analyse acetylated protein levels using an antibody specifically recognizing acetylated-lysine residues. Control cells and dPANK/Fbl-depleted cells were left untreated or treated with pantethine (0.1 mM), with TSA or with TSA and pantethine.
Quantification of the relative levels of acetylation compared to control cells for the indicated band of 55 kD in size and quantification of the relative levels of acetylation for the indicated bands around 17–11 kD.
S2 cells were incubated with 0.5 mM HoPan, and/or with 0.1 mM Pantethine. Whole cell extracts were probed with anti-acetyl-lysine antibody.
dPANK/Fbl depleted cells were left untreated or increasing concentrations of CoA were added to the cell culture medium. Whole cell extracts were probed with an acetyl-lysine antibody.
Quantification of the relative levels of acetylation compared to control cells after addition of various concentrations of CoA for the indicated band of 55 kD in size and quantification of the relative levels of acetylation for the indicated bands around 17–11 kD. Asterisks indicate bands that show a decreased signal in the dPANK/Fbl-depleted or HoPan treated cells. Efficiency of the RNAi treatment was controlled by using an antibody specifically recognizing dPANK/Fbl (Bosveld et al, 2008). As a loading control tubulin was used.
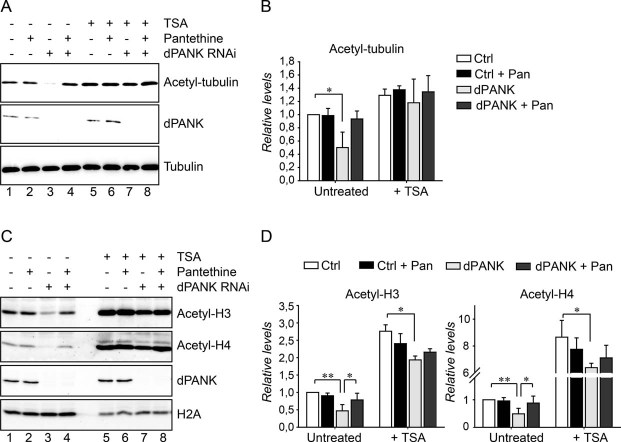
Cell extracts of control cells and dPANK/Fbl-depleted cells (by RNAi) were analysed by Western blot using antibodies specifically recognizing acetylated-tubulin. Efficiency of RNAi was determined by using a dPANK/Fbl antibody and tubulin was used as a loading control. Control cells and dPANK/Fbl-depleted cells were left untreated, treated with pantethine, with TSA or with TSA and pantethine.
Quantification of the relative levels of tubulin acetylation under the conditions presented in A compared to control cells.
Cell extracts of control cells and dPANK/Fbl-depleted cells were analysed using Western blot to determine acetylation levels of specific histones. Specific antibodies were used to detect levels of acetylated histone 3 and acetylated histone 4. Control and dPANK/Fbl-depleted cells were left untreated or were treated with pantethine, with TSA or with TSA and pantethine. The efficiency of the RNAi treatment was investigated by the use of an antibody against dPANK/Fbl. H2A was used as a loading control.
Quantification of the relative levels of histone acetylation under the conditions presented in C compared to control cells.
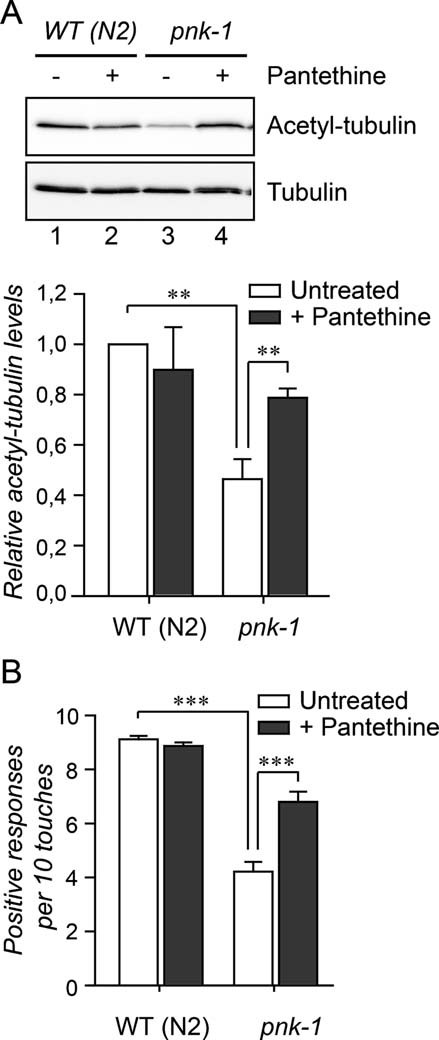
Extracts of staged L4 + 2 WT and pnk-1 mutant (pnk-1) animals were analysed by Western blot using antibodies specifically recognizing acetylated-tubulin. Tubulin was used as a loading control.
Touch responses were scored as previously described (Shida et al, 2010) in WT (animals and in pnk-1 mutants under control conditions and after addition of pantethine to the medium. Error bars indicate SEM.
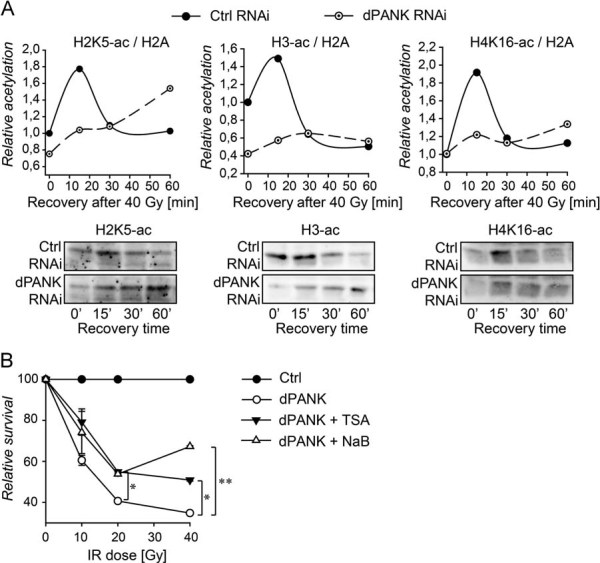
Control cells and dPANK/Fbl-depleted cells were irradiated (40 Gy) and acetylation levels of specific histone tails were determined after various time points. Levels of H3Ac, H4K16 and H2K5 were determined using specific antibodies for each histone or histone tail. The blots and the quantifications are representative of three-independent experiments.
Relative cell survival of control cells and dPANK/Fbl-depleted cells was measured after various doses of irradiation (10, 20 and 40 Gy). Cell survival was also determined after treating the cells with HDAC inhibitors (TSA or NaB; See Fig S4A of Supporting information for acetylation levels). Survival of untreated control cells was set to 100% for every dose of irradiation.
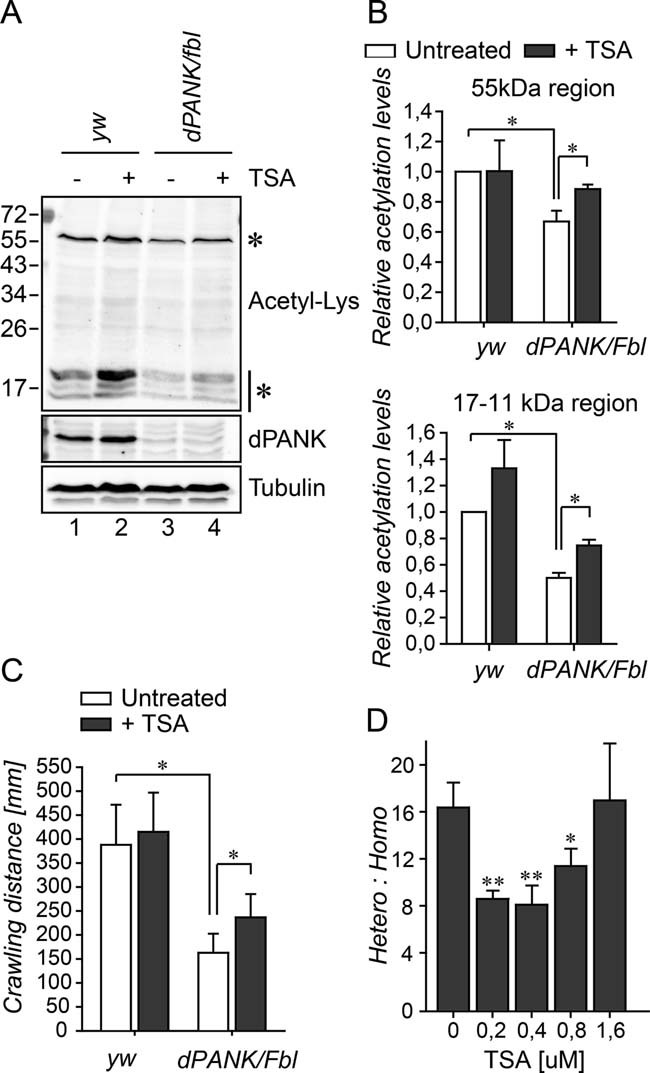
Extracts of WT and dPANK/Fbl homozygous third instar larvae were analysed for their levels of acetylated proteins using acetyl-Lys antibody. Tubulin was used as a loading control and the dPANK antibody was used to demonstrate the reduced expression of dPANK/Fbl in the mutant larvae. Addition of 0.2 µM TSA to the larval food resulted in increased levels of acetylated histones and tubulin.
Quantification of the relative intensity of the 55 kDa band (corresponding to acetyl-tubulin) and <17 kDa bands (corresponding to acetyl-histones) in larvae extracts.
Ability of larvae to crawl a certain distance in 9 min was measured as previously described (Rana et al, 2010). Larval crawling assay was performed in WT larvae and in dPANK/Fbl homozygous mutant larvae untreated or fed with the HDAC inhibitor TSA (0.2 µM).
dPANK/Fbl/TM3 males and females were crossed and various concentrations of TSA were added to the food. The number of homozygous (dPANK/Fbl/dPANK/Fbl) versus heterozygous dPANK/Fbl/TM3 adults which eclosed was counted.
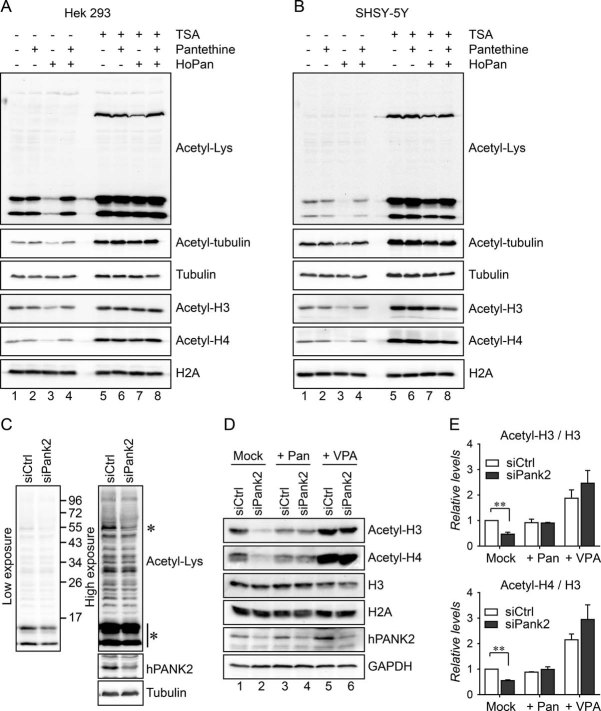
HEK293 cells were left untreated or were treated with HoPan. Control and HoPan treated cells were co-treated with pantethine or TSA. Acetyl-lysine, acetyl-tubulin, acetyl-H3 and acetyl-H4 antibodies were used to analyse acetylation levels of tubulin and histones. Histone H2A and tubulin were used as loading controls.
As in A but human SHSY-5Y cells were used.
HEK293 cells transfected with a control siRNA or with an siRNA against human PANK2 were analysed for the levels of acetylated proteins. Acetyl-lysine antibody was used. Low exposure (left panel) was shown to determine levels of histone acetylation (lowest asterisk), high exposure (right panel) was shown to determine levels of acetyl-tubulin (highest asterisk). Efficiency of RNAi was confirmed using an antibody specifically recognizing human PANK2. Tubulin was used as a loading control.
Control cells and human PANK2 depleted cells were investigated for their levels of acetyl H3 and acetyl H4. Cells were additionally left untreated or were treated with pantethine or VPA. Specific antibodies were used to determine levels of histone H3 and histone H4. The efficiency of human PANK2 RNAi was determined by a specific human PANK2 antibody. H3 and GAPDH were used as loading controls.
Quantification of the relative levels of acetylated histone 3 and acetylated histone 4 compared to control cells.
References
-
- Allis CD, Berger SL, Cote J, Dent S, Jenuwien T, Kouzarides T, Pillus L, Reinberg D, Shi Y, Shiekhattar R, et al. New nomenclature for chromatin-modifying enzymes. Cell. 2007;131:633–636. - PubMed
-
- Biedler JL, Roffler-Tarlov S, Schachner M, Freedman LS. Multiple neurotransmitter synthesis by human neuroblastoma cell lines and clones. Cancer Res. 1978;38:3751–3757. - PubMed
-
- Bosveld F, Rana A, van der Wouden PE, Lemstra W, Ritsema M, Kampinga HH, Sibon OC. De novo CoA biosynthesis is required to maintain DNA integrity during development of the Drosophila nervous system. Hum Mol Genet. 2008;17:2058–2069. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
