

Molecular characterization of disrupted in schizophrenia-1 risk variant S704C reveals the formation of altered oligomeric assembly
- PMID: 21998303
- PMCID: PMC3243500
- DOI: 10.1074/jbc.M111.271593
Molecular characterization of disrupted in schizophrenia-1 risk variant S704C reveals the formation of altered oligomeric assembly
Abstract
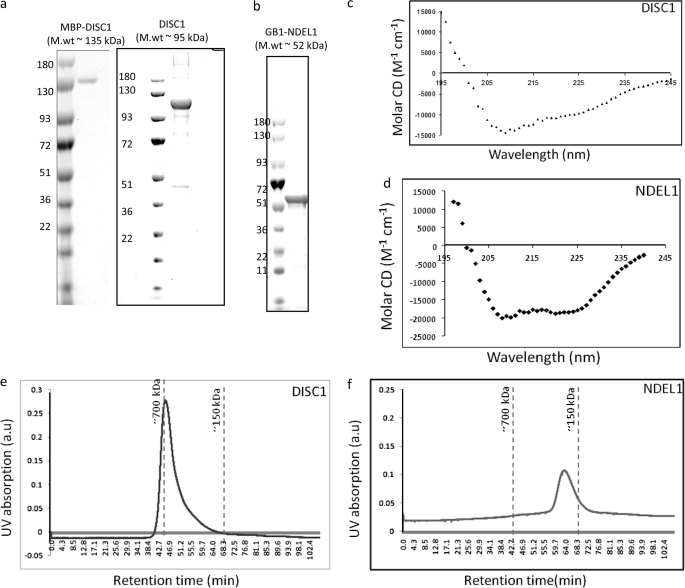
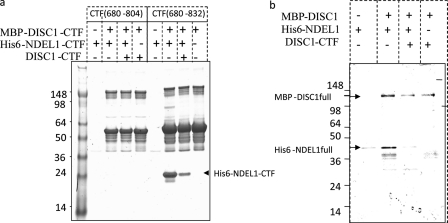
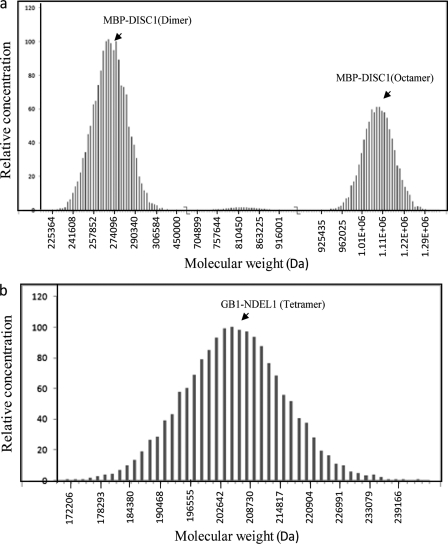
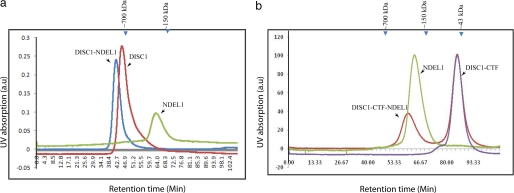
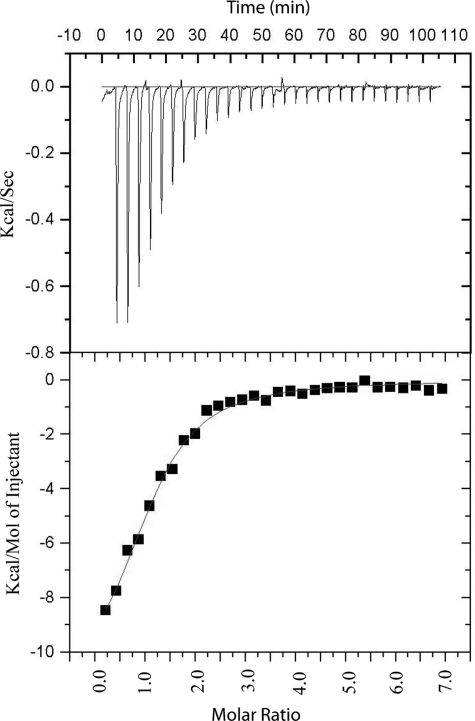
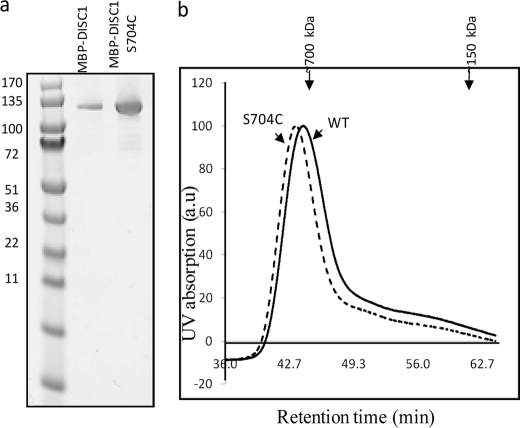
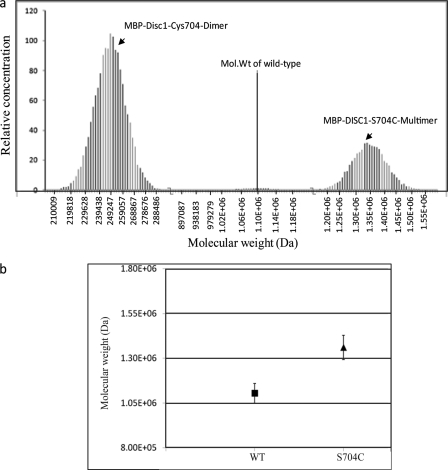
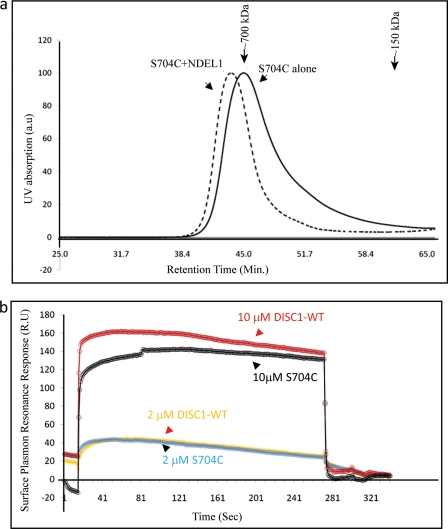
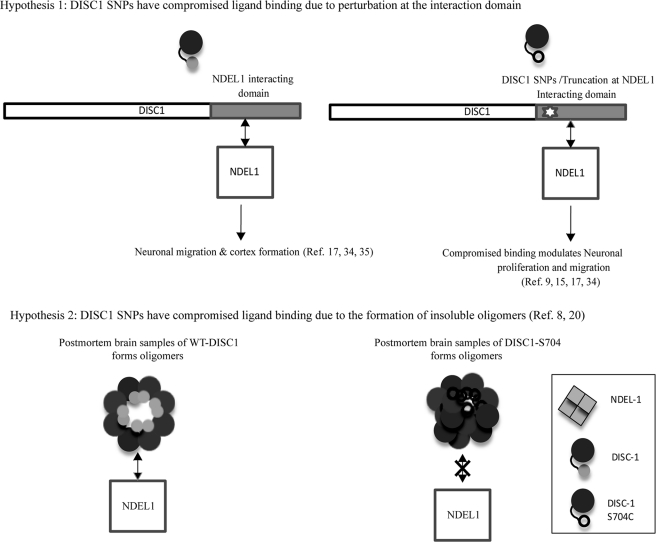
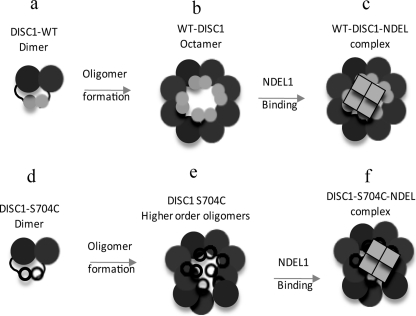
DISC1 (Disrupted in schizophrenia-1) plays essential roles in neuronal proliferation, neuronal migration and axon guidance and has been implicated in schizophrenia and related psychiatric disorders. DISC1 forms a functional complex with nuclear distribution element-like protein-1 (NDEL1), a key component that regulates microtubule organization during cell division and neuronal migration. DISC1 polymorphisms at the binding interface of DISC1-NDEL1 complex have been implicated in schizophrenia. However, it is unknown how schizophrenia risk polymorphisms perturb its interaction with NDEL1 and how they change the inherent biochemical properties of DISC1. Here, we characterize the oligomerization and binding property of DISC1 and its natural schizophrenia risk variant, S704C. Our results show that DISC1 forms octamers via dimers as building blocks and directly interacts with tetramers of NDEL1. The schizophrenia risk variant S704C affects the formation of octamers of DISC1 and exhibits higher-order self-oligomerization. However, the observed formation of new oligomeric species did not influence its binding with NDEL1. These results suggest that the improper oligomeric assembly of DISC1-S704C may underlie the observed phenotypic variation due to the polymorphism.
Figures
References
-
- Mackie S., Millar J. K., Porteous D. J. (2007) Curr. Opin. Neurobiol. 17, 95–102 - PubMed
-
- Millar J. K., Pickard B. S., Mackie S., James R., Christie S., Buchanan S. R., Malloy M. P., Chubb J. E., Huston E., Baillie G. S., Thomson P. A., Hill E. V., Brandon N. J., Rain J. C., Camargo L. M., Whiting P. J., Houslay M. D., Blackwood D. H., Muir W. J., Porteous D. J. (2005) Science 310, 1187–1191 - PubMed
-
- Ross C. A., Margolis R. L., Reading S. A., Pletnikov M., Coyle J. T. (2006) Neuron 52, 139–153 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
