

Evolution of networks for body plan patterning; interplay of modularity, robustness and evolvability
- PMID: 21998573
- PMCID: PMC3188509
- DOI: 10.1371/journal.pcbi.1002208
Evolution of networks for body plan patterning; interplay of modularity, robustness and evolvability
Abstract
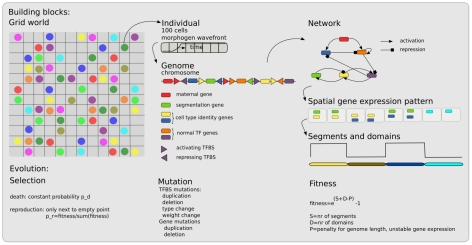
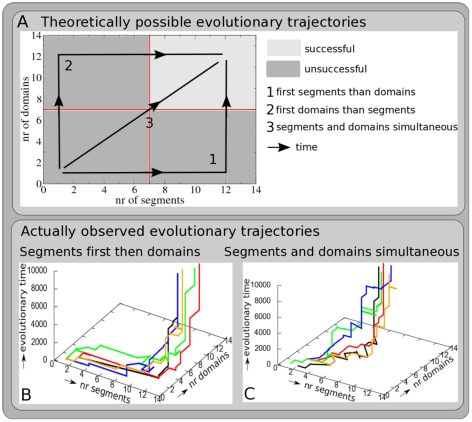
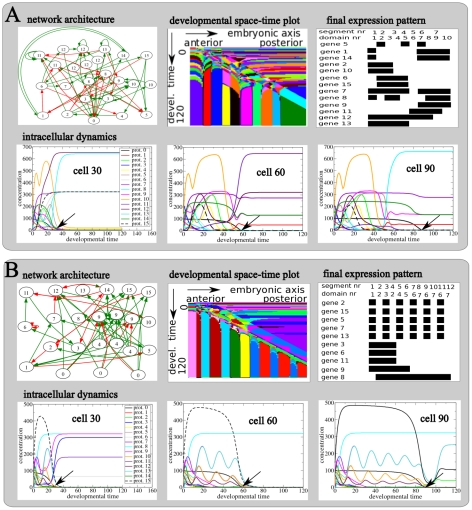
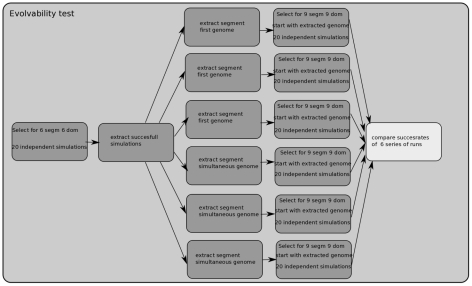
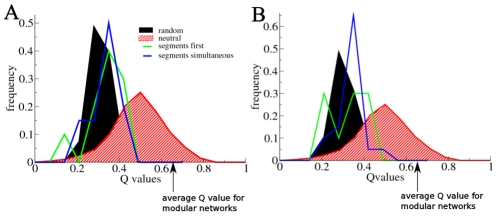
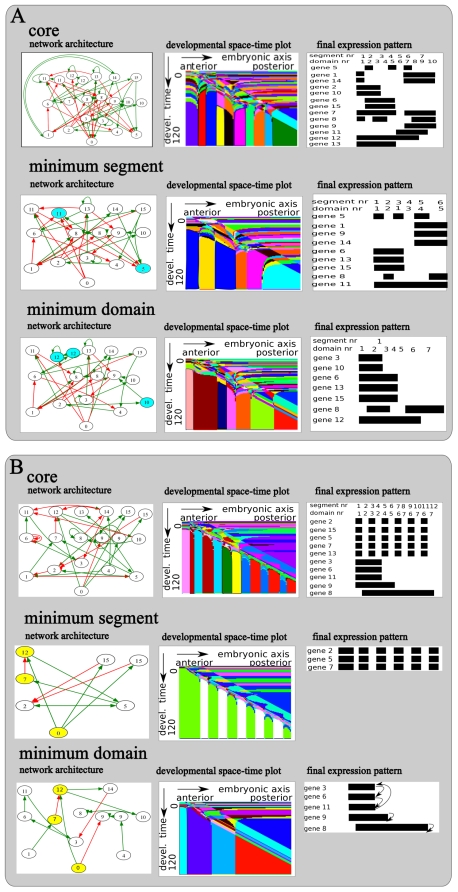
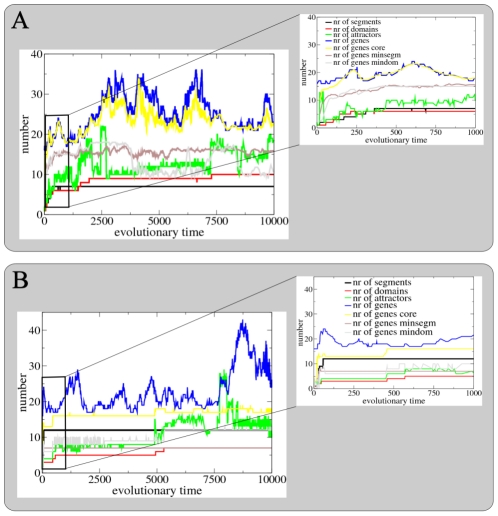
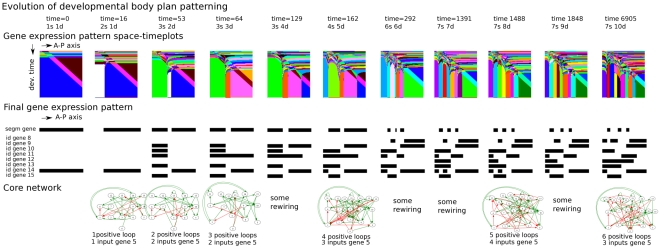
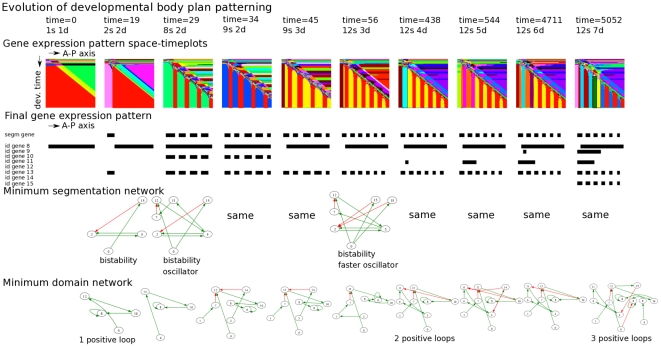
A major goal of evolutionary developmental biology (evo-devo) is to understand how multicellular body plans of increasing complexity have evolved, and how the corresponding developmental programs are genetically encoded. It has been repeatedly argued that key to the evolution of increased body plan complexity is the modularity of the underlying developmental gene regulatory networks (GRNs). This modularity is considered essential for network robustness and evolvability. In our opinion, these ideas, appealing as they may sound, have not been sufficiently tested. Here we use computer simulations to study the evolution of GRNs' underlying body plan patterning. We select for body plan segmentation and differentiation, as these are considered to be major innovations in metazoan evolution. To allow modular networks to evolve, we independently select for segmentation and differentiation. We study both the occurrence and relation of robustness, evolvability and modularity of evolved networks. Interestingly, we observed two distinct evolutionary strategies to evolve a segmented, differentiated body plan. In the first strategy, first segments and then differentiation domains evolve (SF strategy). In the second scenario segments and domains evolve simultaneously (SS strategy). We demonstrate that under indirect selection for robustness the SF strategy becomes dominant. In addition, as a byproduct of this larger robustness, the SF strategy is also more evolvable. Finally, using a combined functional and architectural approach, we determine network modularity. We find that while SS networks generate segments and domains in an integrated manner, SF networks use largely independent modules to produce segments and domains. Surprisingly, we find that widely used, purely architectural methods for determining network modularity completely fail to establish this higher modularity of SF networks. Finally, we observe that, as a free side effect of evolving segmentation and differentiation in combination, we obtained in-silico developmental mechanisms resembling mechanisms used in vertebrate development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Wilkins A. Sinauer; 2002. The Evolution of Developmental Pathways.
-
- Davidson E. Academic Press; 2006. The Regulatory Genome.
-
- Wolpert L. Oxford University Press; 2007. Principles of Development.
-
- Tautz D. Evolution of transcriptional regulation. Curr Opin Genet Dev. 2000;10:575–579. - PubMed
-
- Levine M, Tjian R. Transcription regulation and animal diversity. Nature. 2003;424:147–151. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
