

Noninfectious retrovirus particles drive the APOBEC3/Rfv3 dependent neutralizing antibody response
- PMID: 21998583
- PMCID: PMC3188525
- DOI: 10.1371/journal.ppat.1002284
Noninfectious retrovirus particles drive the APOBEC3/Rfv3 dependent neutralizing antibody response
Abstract
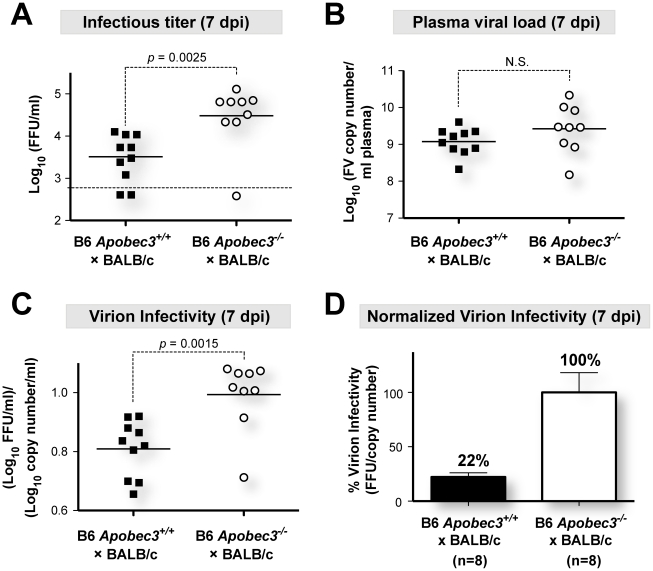
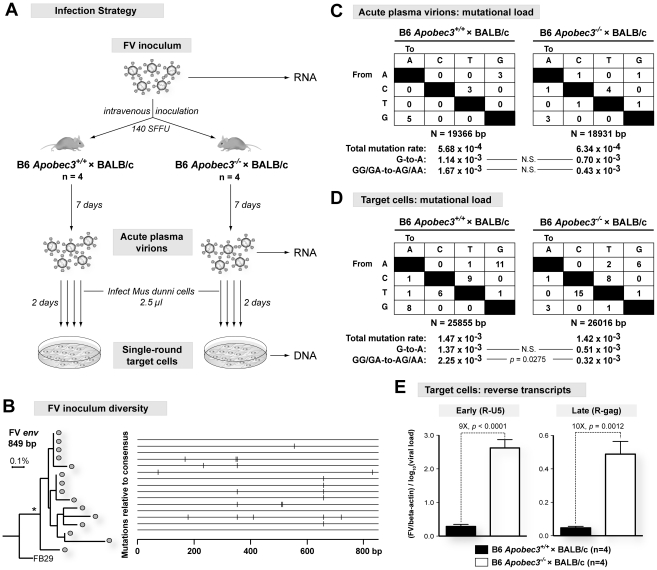
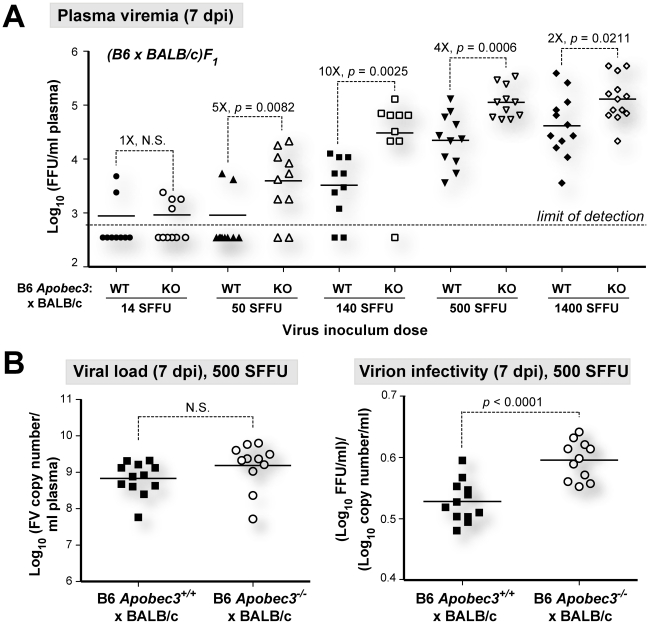
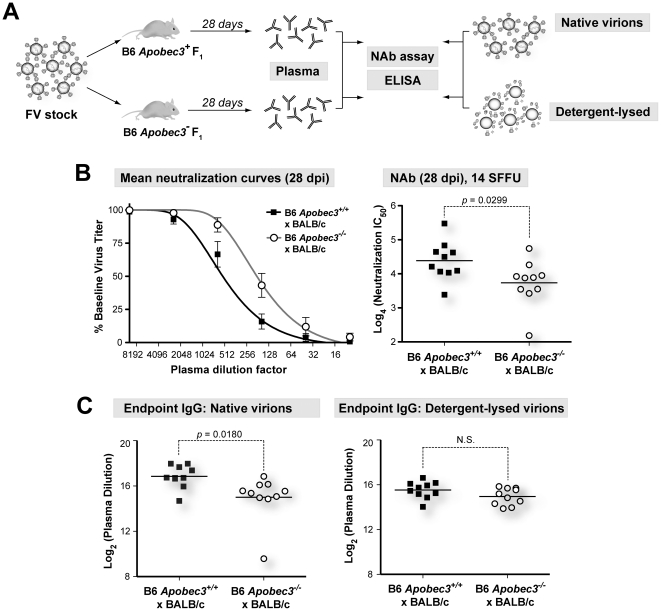
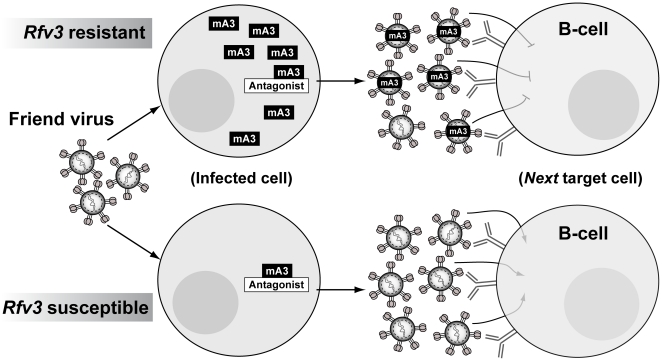
Members of the APOBEC3 family of deoxycytidine deaminases counteract a broad range of retroviruses in vitro through an indirect mechanism that requires virion incorporation and inhibition of reverse transcription and/or hypermutation of minus strand transcripts in the next target cell. The selective advantage to the host of this indirect restriction mechanism remains unclear, but valuable insights may be gained by studying APOBEC3 function in vivo. Apobec3 was previously shown to encode Rfv3, a classical resistance gene that controls the recovery of mice from pathogenic Friend retrovirus (FV) infection by promoting a more potent neutralizing antibody (NAb) response. The underlying mechanism does not involve a direct effect of Apobec3 on B cell function. Here we show that while Apobec3 decreased titers of infectious virus during acute FV infection, plasma viral RNA loads were maintained, indicating substantial release of noninfectious particles in vivo. The lack of plasma virion infectivity was associated with a significant post-entry block during early reverse transcription rather than G-to-A hypermutation. The Apobec3-dependent NAb response correlated with IgG binding titers against native, but not detergent-lysed virions. These findings indicate that innate Apobec3 restriction promotes NAb responses by maintaining high concentrations of virions with native B cell epitopes, but in the context of low virion infectivity. Finally, Apobec3 restriction was found to be saturable in vivo, since increasing FV inoculum doses resulted in decreased Apobec3 inhibition. By analogy, maximizing the release of noninfectious particles by modulating APOBEC3 expression may improve humoral immunity against pathogenic human retroviral infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Stremlau M, Owens CM, Perron MJ, Kiessling M, Autissier P, et al. The cytoplasmic body component TRIM5alpha restricts HIV-1 infection in Old World monkeys. Nature. 2004;427:848–853. - PubMed
-
- Neil SJ, Zang T, Bieniasz PD. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature. 2008;451:425–430. - PubMed
-
- Sheehy AM, Gaddis NC, Choi JD, Malim MH. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature. 2002;418:646–650. - PubMed
-
- Mariani R, Chen D, Schrofelbauer B, Navarro F, Konig R, et al. Species-specific exclusion of APOBEC3G from HIV-1 virions by Vif. Cell. 2003;114:21–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
