

A genetic screen reveals Arabidopsis stomatal and/or apoplastic defenses against Pseudomonas syringae pv. tomato DC3000
- PMID: 21998587
- PMCID: PMC3188540
- DOI: 10.1371/journal.ppat.1002291
A genetic screen reveals Arabidopsis stomatal and/or apoplastic defenses against Pseudomonas syringae pv. tomato DC3000
Abstract
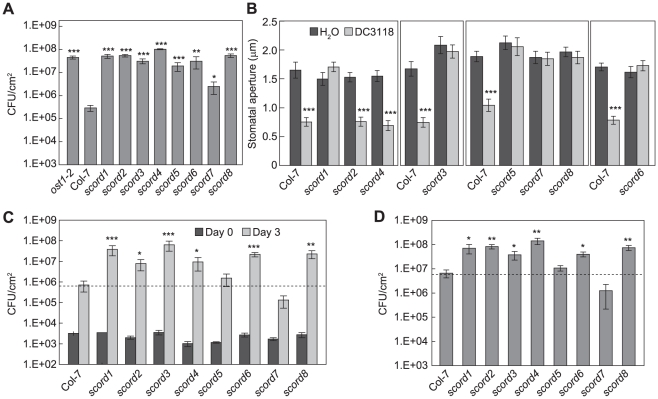
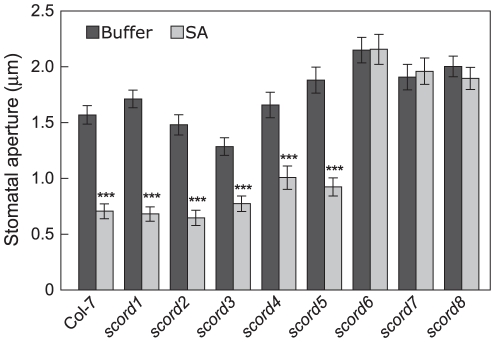
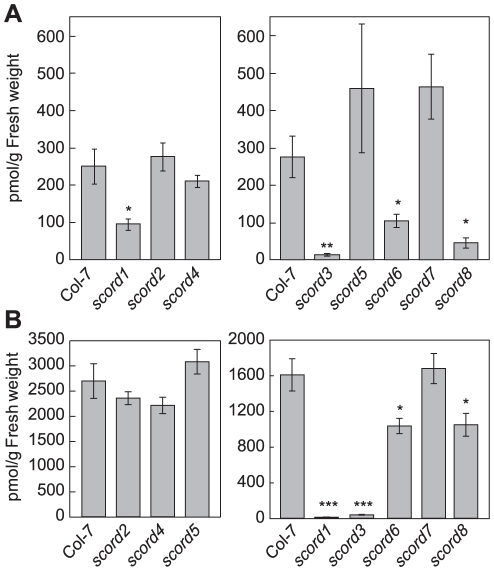
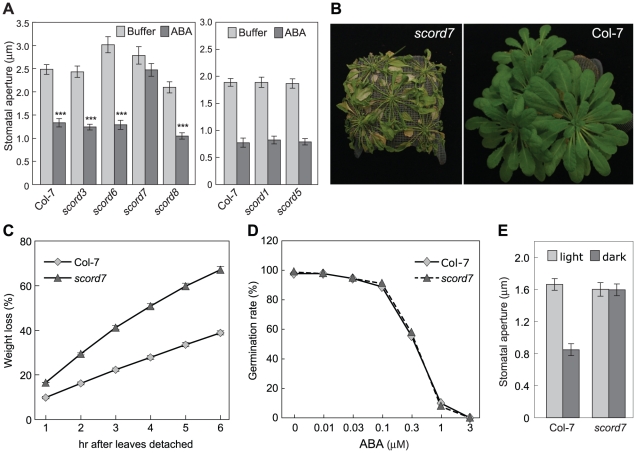
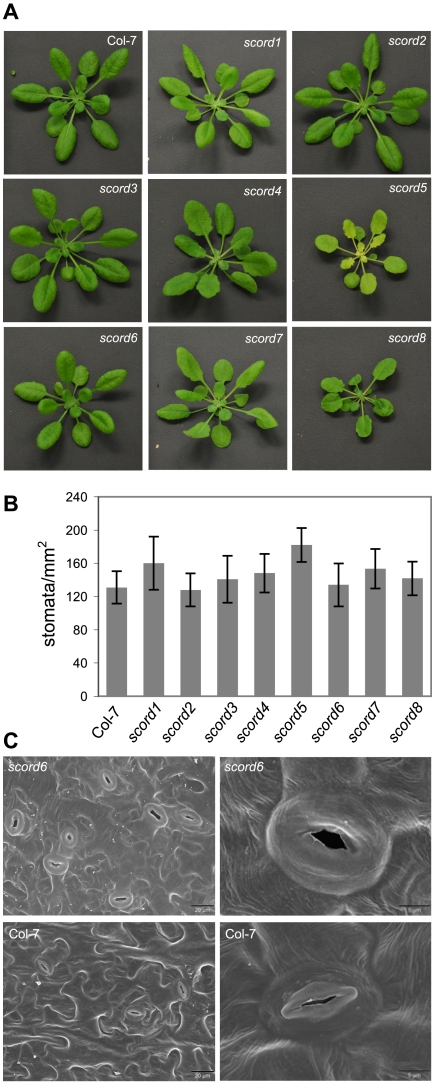
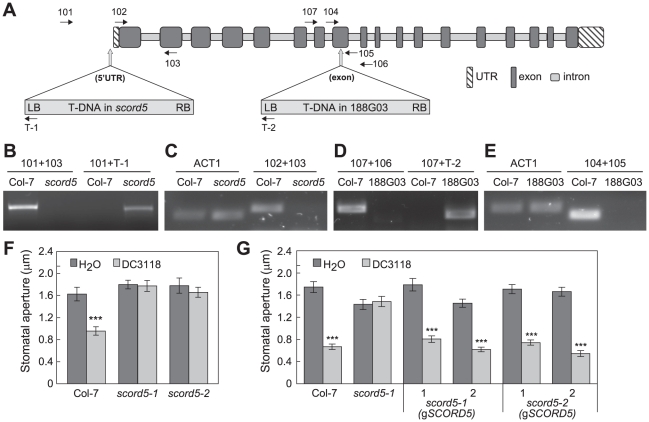
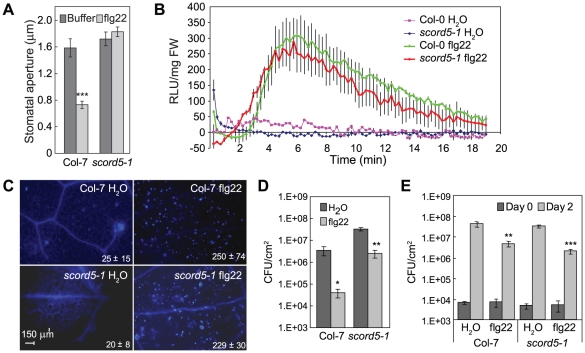
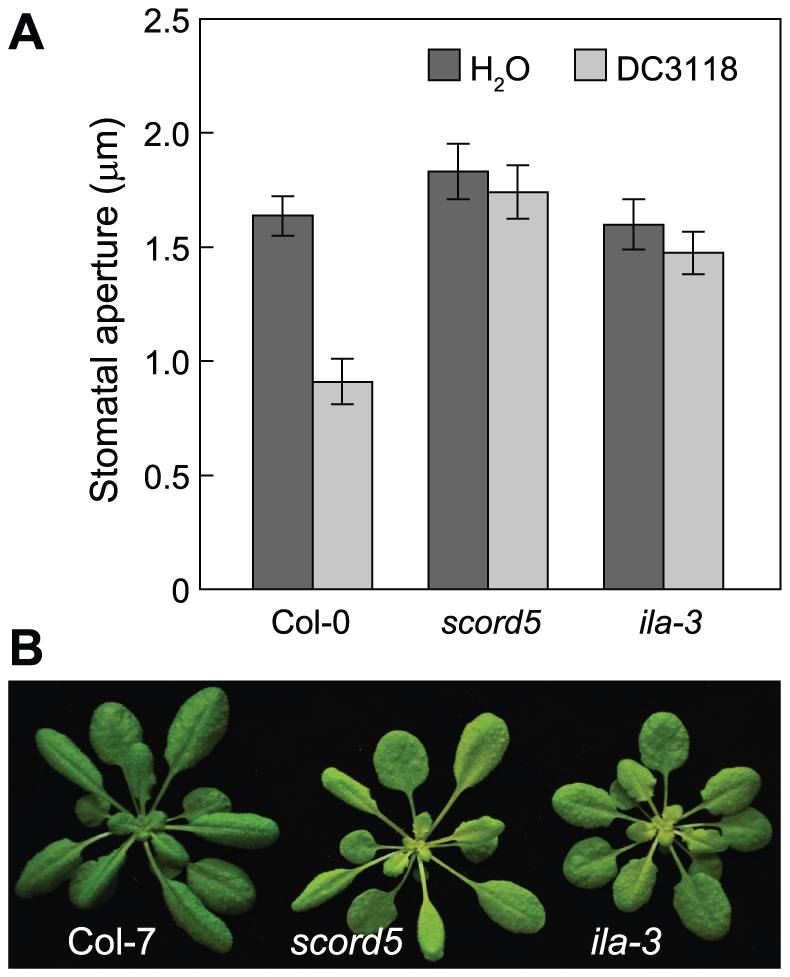
Bacterial infection of plants often begins with colonization of the plant surface, followed by entry into the plant through wounds and natural openings (such as stomata), multiplication in the intercellular space (apoplast) of the infected tissues, and dissemination of bacteria to other plants. Historically, most studies assess bacterial infection based on final outcomes of disease and/or pathogen growth using whole infected tissues; few studies have genetically distinguished the contribution of different host cell types in response to an infection. The phytotoxin coronatine (COR) is produced by several pathovars of Pseudomonas syringae. COR-deficient mutants of P. s. tomato (Pst) DC3000 are severely compromised in virulence, especially when inoculated onto the plant surface. We report here a genetic screen to identify Arabidopsis mutants that could rescue the virulence of COR-deficient mutant bacteria. Among the susceptible to coronatine-deficient Pst DC3000 (scord) mutants were two that were defective in stomatal closure response, two that were defective in apoplast defense, and four that were defective in both stomatal and apoplast defense. Isolation of these three classes of mutants suggests that stomatal and apoplastic defenses are integrated in plants, but are genetically separable, and that COR is important for Pst DC3000 to overcome both stomatal guard cell- and apoplastic mesophyll cell-based defenses. Of the six mutants defective in bacterium-triggered stomatal closure, three are defective in salicylic acid (SA)-induced stomatal closure, but exhibit normal stomatal closure in response to abscisic acid (ABA), and scord7 is compromised in both SA- and ABA-induced stomatal closure. We have cloned SCORD3, which is required for salicylic acid (SA) biosynthesis, and SCORD5, which encodes an ATP-binding cassette (ABC) protein, AtGCN20/AtABCF3, predicted to be involved in stress-associated protein translation control. Identification of SCORD5 begins to implicate an important role of stress-associated protein translation in stomatal guard cell signaling in response to microbe-associated molecular patterns and bacterial infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Preston GM. Pseudomonas syringae pv. tomato: the right pathogen, of the right plant, at the right time. Mol Plant Pathol. 2000;1:263–275. - PubMed
-
- Mitchell RE. Coronatine production by some phytopathogenic Pseudomonads. Physiol Plant Pathol. 1982;20:83–89.
-
- Ma SW, Morris VL, Cuppels DA. Characterization of a DNA region required for production of the phytotoxin coronatine by Pseudomonas syringae pv. tomato. Mol Plant Microbe Interact. 1991;4:69–77.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
